

Калиновскин Г.Н., Шнайдер В.Л.
Житомирский национальный агроэкологический университет
г. Житомир. Украина
Омелянееко Н.Н.
Национальный университет биоресурсов и природоиспользования г. Киев, Украина
Введение. Половой цикл как сложный физиологический нейрогормональный процесс сопровождается соответствующими морфологическими изменениями во всех отделах половых органов и ярко выраженными клиническими признаками наиболее характерными для стадии возбуждения.
Ограниченность информации о состоянии маточных труб и их связи со смежными органами во многих случаях не только осложняет вы-бор способов лечения, но и не позволяет эффективно решать такие важные практические вопросы как осеменение коров, пересадки эмбрионов и профилактики симптоматического бесплодия.
Выяснение изложенных и других вопросов, касающихся морфо-функционального состояния маточных труб в разные стадии полового цикла в норме и при патологии даст возможность разрабатывать и обосновывать эффективные способы не только профилактики, но и лечения бесплодных коров.
Цель работы - изучить морфофункциональное состояние маточных труб коров в стадию уравновешивания полового цикла.
Материал и методика исследований. Материалом для исследования были внутренние половые органы, отобранные от клинически здоровых коров после их забоя на мясокомбинате. При осмотре половых органов определяли соответствие диагнозов в сопроводительных документах на коров с данными анатомических параметров матки и яич-ников. Маточные трубы отпрепаровывали, умеренно растягивали, измеряли их длину и диаметр. Из перешейка, ампулы и воронки высекали кусочки длиной 0,5-0,7 см, из которых по существующей методике
изготавливали гистопрепараты [ 1 ]. Окрашенные гематоксилин-эозином и по Ван-Гизону препараты изучали и фотографировали пользуясь микроскопом МБИ-б.
Результаты исследований и их обсуждение. Маточные трубы представляют собой изогнутые тканевые трубки, состоящие из 17-22 коленец, длиной 0,8-1,2 см. Диаметр трубы состовляет - 0,2-3 мм. Длина маточных труб колеблется в пределах- 11,3-22,6 мм (рис.1).
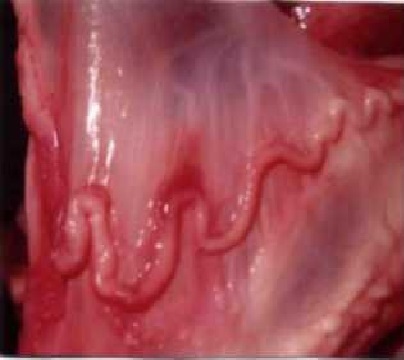
Рис. 1. Маточная труба
нестельной коровы в стадии равновесия полового цикла
Стенка маточной трубы состоит из слизистой, мышечной и серозной оболочек (рис.2), имеет наибольшую толщину в участке перешейка. Щели между складками различной формы и величины. Во всех участках трубы слизистая оболочка выстелена многорядным однослойным цилиндрическим эпителием, поверхность основных складок частично оголена от эпителия (рис. 2). Серозная оболочка в перешейке отсутсвует и мышечный слой прилегает непосредственно к связке трубы. Остов верхушек складок неоднороден, разной толщины, что придает им определенную форму (рис. 2). Ткани связки, прилегающие непосредственно к мышечной оболочки густо васкуляризованные.
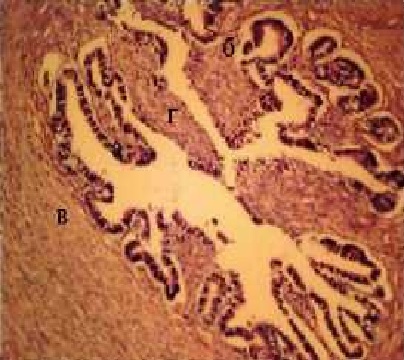
Рис. 2. Фрагмент гистоструктуры перешейка маточной трубы: а - слизистая оболочка; б - складки слизистой оболочки; в - мышечный слой: г - щели между складками
Г.Е.,х400.
Частичное оголение верхушек складок и их стенок свидетельствует о незавершенном восстановлении их выстелки после стадии возбуждения полового цикла.
Диаметр просвета ампулы маточной трубы больший, чем перешей-ка, но толщина стенок меньшая в основном за счет истончения слоев мышечной оболочки. Складки слизистой оболочки ампулы длинее и тоньше, имеющихся в перешейке, щели между ними шире и они полностью закрывают просвет ампулы. Дно просвета ампулы выстелено сплошным шаром однорядного однослойного цилиндрического эпителия, который на поверхности складок становится многорядным, на их верхушках многослойным, на отдельных участках отмечается его скопление и отслоение с оголением стромы складок (рис. 3).
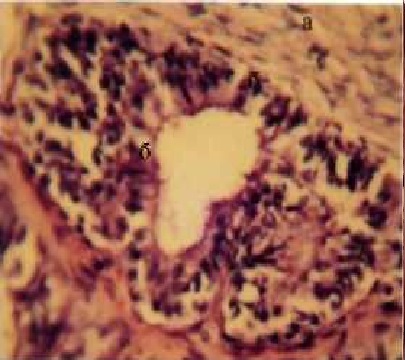
Рис. 3. Фрагмент гистоструктуры ампулы
маточной трубы:
а - стенка:
б - эпителий
Г.Е., х 400
Просвет воронки почти полностью заполнен складками, щели между ними - отслоившимся эпителием, они тонкие и длинные, их поверхность частично оголена от эпителия, верхушки отдельных складок разполокнены и разрушены (рис. 4).
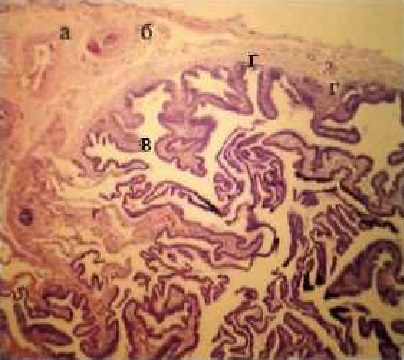
Рис. 4. Фрагмент гистоструктуры воронки
маточной трубы:
а - мышечная оболочка:
б - слизистая оболочка:
в - складки:
г - щели.
Как известно, феномены стадии возбуждения полового цикла про-являются не только соотвествующими клиническими признаками, но и сложными морфологическими изменениями во всех отделах половых органов. Если эти изменения известны и описаны во внешних и внутренних половых органах, прежде всего в матке и яичниках, то их характер в маточных трубах не полностью раскрыт и только в общих чертах упоминается в отдельных работах [2].
Наши исследования свидетельсвуют о том, что морфологические измения во внутренных половых органов коров, ярко выражены в течении стадии возбуждения постепенно угасают, но в маточных трубах они завершается в стадии уравновешывания только частично.
У коровы в перешейке преобладают продольные главные и добавочные складки, которые здесь выше и многочнсленее. Звездообразный просвет, образованный складками, продолжается до брюшного отверствия трубы [3].
Высокие складки изменяются соответственно диаметру трубы. В ампуле между крупными главными складками густо распологаются добавочные складки до третьего-четвертого порядков. Просвет полностью заполняют складки, щели между ними равны диаметру яйцеклетки (100-200 мкм), [7]. Просвети щелеобразные пространства заполнены трубной жидкостью, являющийся секретом эпителия трубы. Складки состоят из эпителиального покрова и соединительнотканной основы, которая в участке ампулы тоньше от других и от эпителиального покрова. Соединительнотканная основа собственного слоя слизи-стой оболочки трубы изменяется соответсвенно стадиям полового цикла. Автор считает, что густая сеть кровеносных и лимфатических капиляров соединительнотканной основы складок в ампулярной и воронковой части трубы придают ей кавернозный вид, что и обеспечивает их эрекцию во время охоты.
Толщина эпителиального слоя складок слизистой оболочки трубы колеблется от 20 до 63,3 мкм, она неодинова в различных участках трубы как на ее дне, так и на поверхности складок: у воронковой части наивысшая, в ампуле несколько ниже, в перешейке - самая низкая. Эти показатели изменяются в течении полового цикла, особенно в ампуле
[4]-
Среди реснитчатых и безреснитчатых клеток эпителиальной выстелки находятся секреторные (железистые) вклинивающиеся или штифтовые и базальные или замещающиеся клетки, которые рассеяны непосредственно над базальною мембраной [3].
Реснитчатые клетки встречаются во всех стадиях полового цикла и при стельности, их свободная поверхность покрыта микроворсниками и ресничками [6]. Наибольшая длина ресничек наблюдается во время охоты и после течки.
В стадии уравновешываня, перед началом стадии возбуждения (передтечковый период) морфологическая структура маточных труб характеризуется равномерной высотой поверхности слизистой оболочки, выстеленой епителием одинаковой высоты, появленим в субнуклеарной зоне клеток, зернышек различной формы, окрашенных в темно-синий цвет со смещением ядра в апикальную часть клетки, удлинением эпителиальных клеток и появлением глыбок под их ядрами. Между эпителиальными клетками появляются клетки с пикнотическими ядрами, отмечается вазодилятация и отек собственно слизистой оболочки.
Во время яркого проявления феноменов стадии возбуждения полового цикла высота эпителиальной выстелки увеличивается, особенно в ампуле, из цитоплазмы вокруг ядер исчезают зернышки и глыбчатые образования, усиливается вазадилитация слизистой оболочки [5].
В стадию торможения полового цикла наступает постепенное угасание возникших изменений, уменьшается отек слизистой и высота эпителиальных клеток, появляется вакулизация цитоплазмы, базальное смещение ядер. Все изменения маточной трубы соответсвуют секреторной активности половых гормонов [3].
Отделение от поверхности слизистой оболочки эпителального слоя
цитоплазматичесюгх клеток, которые считали как признак голокриновой секреции, не нашло подтверждения, так как собственно секреторным процесом является лишь выделение секреторных зернышек, наблюдаемое до вознокновения цитоплазматических выростов, что свойственно для мероклиневого типа секреции [6].
Заключение. Проведенные исследования свидетельсвуют о том, что морфологические измения во внутренных половых органов коров, ярко выражены в течении стадии возбуждения постепенно угасают, но в маточных трубах они завершается в стадии уравновешывания только частично.
ЛИТЕРАТУРА
1. Меркулов Г.А. Курс патологистоллогической техники / Г.А. Меркулов. - Издательство «Медицина» Ленинградское отделение, Изд.5., 1969., - 423 с.
2. Милованов В.К. Иммунология воспроизведения / В.К. Милованов, И.И. Соколовская//М., колос, Тарту. 1967.- 194 с.
3. Техвер Ю.М. Гистология мочеполовых органов и молочной железы домашних животных /Ю.М. Техвер.-Ч.П., 1968.-С. 188-203.
4. Чхартишвили Т.Е. Закладка и дальнейшее развитие яйцеводов сельскохозяйственных животных (коров, буйволов, овец, свиней и прочих) и изменения строение слизистой оболочки во время течки и беременности / Т.Е. Чхартишвили // Автореф. дисс, Тбилисси. 1955
5. Lombard T.F.; Morgan В. С. The morfphology of the oviduct of virgin heifrs in relation to the estrous cucle. J. Morphology. 1950. - 86.1.
6. Nilson O. Electron microscopy of the Fallopian tube epithelium of rabbits in oestrus. Exptl. Cell Res. 1958. - 14.2.
7. Schilling E. Untersuchungen uber clen Bau. Den Bau und die Arbeitsweise des Eeleiteis vom Schaf und Ring. Zbl. Veteiinarmediciu. 1962. - 9. 9;839.
СОСТОЯНИЕ МАТОЧНЫХ ТРУБ КОРОВ В СТАДИИ УРАВНОВЕШИВАНИЯ ПОЛОВОГО ЦИКЛА
- Подробности
- Категория: Воспроизводство крс
- Вы здесь:
-
Главная

-
КРС
-
Воспроизводство
- СОСТОЯНИЕ МАТОЧНЫХ ТРУБ КОРОВ В СТАДИИ УРАВНОВЕШИВАНИЯ ПОЛОВОГО ЦИКЛА