
С.Г. КУЗНЕЦОВ, Т.С. КУЗНЕЦОВА
ЗАО "Витасоль" А.С. КУЗНЕЦОВ ООО "Фидленд"
В современных условиях контроль за обеспеченностью животных питательными, минеральными и биологически активными веществами имеет особенно важное значение, так как заболевания, связанные с их недостаточностью, дисбалансом и токсичностью, получили широкое распространение. Кроме того, у животных наблюдают новые формы недостаточности и токсикозов: остеохондрозы, остеодистрофии. кетозы. артрозы, слабость конечностей, мышечные дистрофии молодняка, послеродовые осложнения у высокопродуктивных коров (гипокалыдиемия, гипофосфатемия, анемия), снижение воспроизводительной способности, стрессустойчивости и неспецифической резистентности организма, отравление экотоксикантами, изменение поведения, образование камней в моче- и желчевыводящих путях, нарушение функции щитовидной железы.
В экспериментальных условиях недостаточность витаминов, минеральных элементов и других веществ имеет характерное клиническое проявление. Однако на практике у животных наблюдают стертые и осложненные формы, что затрудняет постановку диагноза. Более того, нарушения обмена веществ часто протекают без каких-либо клинических признаков. Например, недостаточное или избыточное обеспечение животных минеральными веществами ведет к снижению использования питательных веществ корма, продуктивности, качества продукции, воспроизводительной способности и устойчивости к болезням. Такую форму патологии можно распознать только при биохимических исследованиях. При выявлении недостаточности или токсичности минерального элемента нужно учитывать биохимические характеристики крови, молока, органов, тканей, экскретов и волосяного покрова; содержание минеральных веществ в почве, воде и кормах; клинические признаки; уровень продуктивности; ответную реакцию организма на минеральные добавки.
При несбалансированном кормлении у животных ухудшаются аппетит, использование питательных веществ корма, снижаются воспроизводительная функция и продуктивность, нарушается структура волосяного покрова. Однако это может быть следствием недостатка или избытка многих факторов. В том случае, когда элемент тесно связан с одним органом
или функцией организма (например, йод с щитовидной железой), клиническая картина бывает однообразной и довольно специфичной. Однако нарушение функции щитовидной железы могут вызывать и гойтрогенные вещества, содержащиеся в рапсе, капусте, сурепке, льняном шроте, клевере белом, соевых бобах, горохе и др., а также некоторые лекарства. Недостаточность таких элементов, как медь и цинк, проявляется весьма разнообразно, что связано с участием их в биосинтезе многих ферментов. Дефицит элементов может быть вторичным или комплексным, возможно также одновременное проявление недостатка одного элемента и избытка другого: соответственно Си и 2п, Си и Mo, Cd и 2п, Мп и Fe, Си и Pb.
Известно, что определенные метаболические процессы могут нарушаться как при недостатке, так и избытке многих элементов. Например, аналогичные или очень близкие поражения скелета бывают при недостатке Са, Р, Си, Мп, Zn, Si, витаминов А и D, а также при избытке Мо, F, Sr, витамина D. Анемию может вызывать недостаток Fe, Си, Со, некоторых витаминов или избыток в рационе Мп, Mo, Zn, Си, Pb, Se. Снижение и извращение аппетита отмечают при дефиците протеина, Са, Р, Na, Со, Си, Zn и при избытке многих элементов, в связи с этим при оценке статуса питательных, минеральных и биологически активных веществ основное внимание должно быть уделено оперативному своевременному выявлению субклинических стадий их недостаточности, токсикоза и организации профилактических мероприятий.
Рассмотрим вклад биохимических показателей крови, которые наиболее часто исследуются в ветеринарной практике, в оценку обеспеченности организма питательными, минеральными и биологически активными веществами и состояния здоровья животных (см. таблицу).
Уровень кальция в крови здоровых животных зависит от содержания в рационе Са, Р, Мд, витамина D, состояния гормональной и пищеварительной систем, почек и других органов. Содержание кальция в крови падает при длительном дефиците его в рационе, плохом усвоении вследствие недостатка витамина D и паратгормона. Гипокальциемия сопровождает остеодистрофию, рахит, послеродовой парез, гипофункцию околощитовидных желез, она возможна при нефрозе и нефрите. При субклини ческой форме недостаточности кальция в плазме (сыворотке) крови снижается концентрация кальция (ниже 8,0 мг%), повышается активность щелочной фосфатазы (ЩФ) и содержание неорганического фосфора и магния; в моче увеличивается концентрация фосфора, магния, оксипролина; а в костной ткани возрастает активность ЩФ (в 2 - 3 раза) и уменьшаются содержание золы, Са, Р, Мд, плотность и прочность костей.
При избытке кальция в корме (2 % на сухое вещество) концентрация этого элемента в плазме крови возрастает в 1,5 раза, а неорганического фосфора снижается в 3 раза, активность ЩФ не изменяется. Повышение Са в крови может быть при передозировке витамина D. гиперфункции паращитовидных желез.
Все виды обмена веществ в организме неразрывно связаны с превращением фосфорной кислоты. Уровень фосфора в крови завис**7 от тех же факторов, что и содержание кальцииПри дефиците его в рационе в плазме крез* снижается концентрация неорганического фосфора (менее 4 мг%), повышаются активное* ЩФ, содержание Мд и Са; в моче уменьшается концентрация фосфора и возрастает Мд, Са = 5 - 10 раз), оксипролина; в костной ткани падэе содержание Р, Са, Мд, золы, изменяется ее плотность и прочность.
Схема дифференциальной диагностики субклинической формы минеральной недостаточности животных
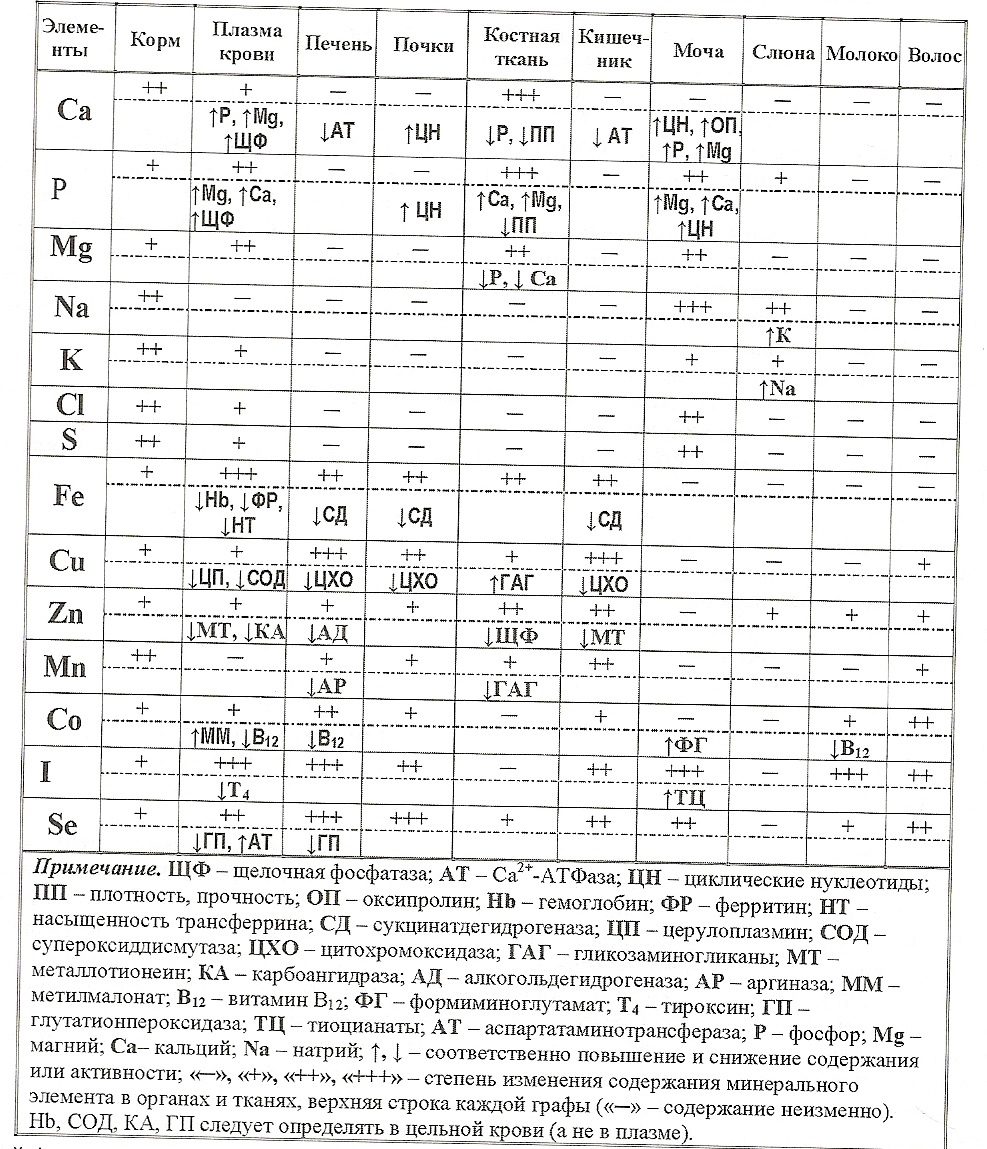
При длительном недостатке наблюдают железодефицитное состояние: нормальная концентрация гемоглобина и снижение запасов железа в тканях. У молодняка при дефиците Fe снижается концентрация его в плазме крови (<65 мкг% ), а также гемоглобина (<80 г/л), гематокрита (<30 %), насыщенности трансферрина (<25 %), эритроцитов (<4,0 -1012/л) и возрастает общая железосвязывающая способность плазмы крови (> 100 мкмоль/л). Из этих показателей наиболее надежными для диагностики ранних форм недостаточности данного элемента считаются содержание его и ферритина в плазме крови, насыщенность трансферрина, концентрация гемоглобина. При избытке железа в рационе (1 г/кг сухого вещества) содержание его возрастает в кишечнике, почках, селезенке и печени.
Субклиническая форма недостаточности меди сопровождается снижением активности церулоплазмина (в 10 - 15 раз) и содержания меди в плазме крови (менее 60 мкг%), а также количества гемоглобина и эритроцитов. Доказано, что активность Си- и Zn-содержащей супероксиддисмутазы в эритроцитах - наиболее надежный индикатор статуса меди у животных, так как она в меньшей степени, чем активность церулоплазмина, зависит от вторичных факторов и содержания меди в плазме крови. Нарушение защитного антимикробного механизма в нейтрофилах - самое раннее проявление недостаточности меди. Избыток ее в рационе приводит к накоплению элемента в печени, почках, стенке кишечника; при этом активность церулоплазмина и содержание Си в плазме крови существенно не изменяются, но развивается гипоцинкемия. Избыток меди и церулоплазмина в крови наблюдают при заболеваниях печени, лейкемии, инфекционных болезнях, при беременности.
При дефиците цинка у коров выявляют умеренное снижение его концентрации в плазме крови (ниже 50 мкг%), костной ткани, семенниках, печени, почках, поджелудочной железе, стенке кишечника, сердце, волосяном покрове, слюне. При избытке элемента отмечают повышение его содержания в основном в этих же тканях и органах. Около 80 % от общего количества цинка в крови приходится на долю эритроцитов, где он соединен главным образом с карбоангидразой; в связи с этим гемолиз повышает содержание цинка в плазме. Концентрация этого элемента в сыворотке крови на 16 % выше, чем в плазме. В практических условиях содержание цинка в плазме крови зависит от вторичных факторов (воспаления, инфекции, опухоли, стрессы, избыток Са, Р, Си, Cd, РЬ), возраста и физиологического состояния животных, их генетических особенностей. Клиниче-
в рационе, плохом усвоении или расстройствах желудочно-кишечного тракта, при недостатке витамина D и паратгормона, остеодисторофии, рахите, уровской болезни, пеллагре.
Избыток фосфора в корме приводит к увеличению в плазме крови количества этого элемента, снижению концентрации магния и не изменяет содержания в ней кальция и активности ЩФ. В моче наблюдают аналогичную картину. Гиперфосфатемию обнаруживают при уменьшении секреции паратгормона, при сердечной недостаточности, кетозе, передозировке витамина D, нефритах, нефрозах, токсикозах, мышечном перенапряжении.
При гиповитаминозе D в плазме крови возрастает активность ЩФ и снижается концентрация Са, Р, Мд. Избыток витамина D приводит к значительному увеличению содержания Са и Р в плазме крови.
Дефицит магния в рационе сопровождается уменьшением концентрации этого элемента в плазме крови (менее 1,7 мг%), моче и костной ткани, а также умеренным снижением содержания Р и Мд в костях, без существенного изменения концентрации Са и P и активности ЩФ. Снижение содержания магния в калия и азота в рационе.
В практических условиях недостаточность натрия, калия, хлора, серы встречается редко, так как в рационы обычно всегда добавляют соль и серосодержащие аминокислоты. Однако у высокопродуктивных животных (особенно в пастбищный период) может наблюдаться недостаточность Na и S. Дефицит хлора даже без добавок поваренной соли - крайне редкое явление, а недостаточность калия может быть у жвачных при высококонцентратном типе кормления, поскольку зерновые корма бедны этим элементом. При дефиците К, S, CI снижается концентрация их в плазме крови и моче. Избыток Na, CI, К, S выделяется в основном с мочой.
Электролиты играют важную роль в поддержании кислотно-щелочного баланса (КЩБ) в организме. Снижение резервной щелочности в крови ниже 40 об.% СОг свидетельствует о сдвиге КЩБ в сторону ацидоза. Метаболическое проявление недостаточности Zn может возникнуть до существенного снижения его содержания в крови и тканях. Эти же факторы способствуют усилению экскреции его с мочой. В наших опытах при дефиците цинка у коров концентрация его в моче не изменялась, а в фекалиях резко снижалась. К настоящему времени только в печени обнаружено около 100 Zn-содержащих ферментов, однако ни один из них пока не может быть использован в качестве надежного критерия статуса Zn в организме.
В эксперименте при недостаточности цинка снижается активность ЩФ в плазме крови, печени, костях. Уменьшение ее в плазме крови еще в большей мере зависит от вторичных факторов, чем содержание цинка. Даже при избытке цинка в корме уровень ЩФ снижается. Активность фермента в плазме крови повышается при дефиците витамина D, рахите, остеодистрофии, гепатите, циррозе печени, а также в конце стельности из-за быстрого роста костной ткани плода. У телят активность ЩФ в 5 - 15 раз выше, чем у взрослых коров.
Содержание марганца в тканях мало изменяется как при недостатке, так и избытке его в рационе. Пока не обнаружены депонирующие его органы. Тем не менее, при его недостаточности содержание элемента умеренно снижается в плазме крови (<2,2 мкг%), печени (<6 мг/кг сухого вещества), почках (<4 мг/кг сухого вещества), волосе (<6 мг/кг сухого вещества), коже, кишечнике, головном мозге, желчи. Концентрация марганца в крови очень низка и определять ее технически трудно. При токсикозе содержание элемента возрастает в стенке кишечника, почках, коже, волосе, желчи и фекалиях. При дефиците этого элемента снижается активность аргиназы в печени, галактозилтрансферазы в костях, изоцитрат- и пируватдегидрогеназы в почках и мышцах, а также концентрация и скорость сульфатирования гликозаминогликанов в хрящевой и костной тканях.
Недостаточность кобальта часто обнаруживают у жвачных в пастбищный период. При дефиците этого элемента снижается его концентрация в плазме крови (<0,6 мкг%) и печени (<0,1 мг/кг сухого вещества), но ввиду низкого содержания кобальта в тканях и биологических жидкостях эти данные имеют небольшую диагностическую ценность. Более надежным показателем является концентрация витамина В12 в плазме крови (у крупного рогатого скота в норме 250 - 600 нг/л), печени и молоке. Однако применяемый в настоящее время микробиологический метод определения витамина В,2 весьма трудоемок и не дает возможности разделить кобаламин на активные и неактивные формы. На величину корреляции между содержанием кобаламина в крови и печени влияют кроме содержания кобальта структура рациона и уровень в нем энергии. При недостаточности кобальта у жвачных резко снижается активность метилмалонил-КоА-мутазы и метионинсинтетазы, что приводит к накоплению в крови метилмалоновой и формиминоглутаминовой кислот и к повышенной экскреции их с мочой. Определение в плазме крови метилмалоната (в норме <4,6 мкмоль/л) и в моче формиминоглутамата (в норме < 100 мкмоль/л) позволяет выявлять субклиническую форму недостаточности кобальта. При избытке в рационе кобальт накапливается в печени и почках.
При недостатке и избытке селена в рационе быстро изменяется его содержание в плазме крови, эритроцитах, сердце, скелетных мышцах, печени, почках, костной ткани, легких, поджелудочной железе, коже, волосе, селезенке, молоке, моче, фекалиях. В плазме крови и особенно в волосе концентрация селена при дефиците элемента снижается медленнее, чем в тканях. Кроме того, определять селен в плазме крови методически трудно из-за низкой его концентрации. Поэтому наиболее надежным прижизненным критерием недостаточности селена является активность Se-зависимой глутатионпероксидазы в цельной крови. Активность данного фермента в плазме крови животных также можно использовать для этой цели, хотя в плазме только около 10 % элемента связано с глутатионпероксидазой и активность ее в 25 - 100 раз ниже, чем в эритроцитах. При дефиците селена в плазме крови возрастает активность аспартатаминотрансферазы и креатинкиназы в 2 раза и более, что является следствием патологии мышечной ткани. Активность глутатионпероксидазы повышается с возрастом животных и может снижаться при дефиците железа и меди.
Многие органы, ткани и экскреты (щитовидная железа, легкие, поджелудочная железа, плазма крови, печень, селезенка, почки, мышцы, волос, молоко, моча) хорошо отражают статус йода в организме. Определение содержания общего, белково-связанного, свободного, бутанолэкстрагируемого йода (БЭЙ) и тироксина в плазме крови позволяет выявитг недостаток этого элемента; в то же время при избытке йода в рационе концентрация гормонов щитовидной железы и БЭЙ не увеличивается. При вторичной недостаточности йода вызванной гойтрогенными веществами, содержание БЭЙ в крови также не изменяется, однако в моче при этом резко повышается уровеш тиоцианатов. У лактирующих животных концентрация йода в молоке - наиболее яркий индикатор обеспеченности организма этим элеметом. Волос, особенно черный, также хорошо отражает статус йода, однако следует учитывать возможность его загрязнения, в том числе и за счет поглощения элемента из воздуха.
В природе известно более 50 каротиноидов, способных превращаться в организме в витамин А. Из них наиболее распространенный и эффективный - бета-каротин, из одной молекулы которого в организме животных образуется 2 молекулы витамина А. В растениях также содержатся зеленый пигмент хлорофилл, желтый - ксантофилл, красный -ликопин и др. Бета-каротин кроме участия в синтезе витамина А выполняет очень важные функции в организме. Он является природным антиоксидантом, повышает сопротивляемость организма к различным заболеваниям, улучшает воспроизводительную функцию, кроветворение и устойчивость организма к раковым опухолям, снижает риск сердечно-сосудистых заболеваний, воспаления слизистых оболочек, регулирует иммунные реакции, усиливает обмен энергии. Обнаружена прямая взаимосвязь между содержанием бета-каротина в организме и воспроизводительной способностью животных.
Обеспечить животных бета-каротином в зимне-стойловый период содержания практически невозможно из-за быстрого его разрушения в процессе заготовки и хранения кормов. Под действием света и высоких температур бета-каротин переходит в менее активные формы. Потребность в бета-каротине возрастает при избытке в рационе энергии, протеина, клетчатки, сульфатов, железа, тяжелых металлов, нитратов, нитритов, микотоксинов, прогорклых жиров, сапонинов, фитоэстрогенов, при воспалении кишечника, воздействии различных стресс-факторов, при лечении кокцидиостатиками и некоторыми другими ветпрепаратами. Повышают усвоение бета-каротина витамины Е и В,2, оптимальные дозы фосфора, цинка, селена, кобальта и жира, углеводы.
Концентрация каротина в крови имеет значительные сезонные колебания (0,4 -1,0 мг% в стойловый период и более 1 мг% - в пастбищный). Уровень его снижается в крови при дефиците его в корме, плохом усвоении из-за влияния многих вторичных факторов, гепатитах, гепатозах, недостатке в рационе белка, жиров, легкоусвояемых углеводов, при различных токсикозах, включая нитратные. Уровень каротина в сыворотке (плазме) крови при хранении образцов снижается, что следует учитывать при проведении анализов и интерпретации полученных результатов (в конце пастбищного периода часто обнаруживают следы каротина в крови и рекомендуют вводить в рацион его препараты).
Доказано, что усваивается только 1/3 - 1/4 часть каротина и около 1 /7 части его превращается в витамин А. 25 - 50 % последнего переходит в печень. Полноценное белковое питание, хорошая обеспеченность витамином В12 и антиоксидантами повышают эффективность превращения каротина в витамин А. Концентрация их в крови имеет значительные сезонные колебания (25 - 80 мкг% в стойловый период и 40 -150 мкг% - в пастбищный). Снижение витамина А в крови и печени ниже соответственно 10 и 50 мкг/г сопровождается развитием клинических признаков гиповитаминоза А. Уменьшение концентрации витамина А в крови, печени, молозиве и молоке отмечают при недостатке каротина и витамина А в кормах, плохом усвоении их вследствие хронических заболеваний желудочно-кишечного тракта и печени.
Снижение уровня общего белка в сыворотке крови (ниже 60 г/л) отмечают при длительном недокорме животных, неполноценном кормлении, остеодистрофии, хронических заболеваниях пищеварительной системы, почек и печени.
Мочевина - основной конечный продукт азотистого обмена , синтезируется в печени и стенке рубца, выделяется главным образом почками. Концентрация ее в крови здоровых животных составляет 20 - 40 мг%, или 3,3 - 6,7 мкмоль/л. Значительное повышение содержания мочевины в крови наблюдают при почечной недостаточности и различных заболеваниях почек, а также при скармливании животным больших количеств зеленых бобовых кормов и передозировках синтетических азотистых веществ (мочевины и др.). Уменьшение содержания мочевины в крови бывает при длительном белковом недокорме, при нарушении мочевинообразовательной функции печени. Такое явление часто встречают у коров с дистрофией печени после переболевания их кетозом.
У коров сразу после отела отмечают быстрый рост молочной продуктивности при дефиците питательных веществ и энергии, которые они получают с кормом. Это, во-первых, связано с пониженным потреблением кормов, а во-вторых, с недостаточным содержанием в них энергии и белка. На этом фоне возможно развитие синдрома ожирения печени, ухудшение воспроизводства, развитие кетоза. Профилактике последнего следует уделять особое внимание, так как заболевание приводит к снижению потребления корма, значительному ухудшению общего состояния животного, ослаблению репродуктивной функции и учащению случаев возникновения хромоты. Развитию кетоза способствует дача в начале лактации большого количества быстро ферментируемых концентратов, что приводит к снижению рН в рубце и является причиной уменьшения содержания жира в молоке и общего удоя.
Кетоновые тела (бета-оксимасляная кислота, ацетоуксусная кислота, ацетон) - промежуточные продукты обмена белков, жиров и углеводов. Повышение уровня кетоновых тел в крови, моче и молоке свидетельствует о нарушении обмена веществ. Стойкая кетонемия встречается у коров при острой и подострой формах кетоза. При этом соотношение кетоновых тел меняется в сторону увеличения ацетона и ацетоацетата.
При остром кетозе обнаруживают ацетоновые тела - в моче до 100 - 500 мг% при норме 5 - 10 мг%, в молоке до 20 - 80 при норме до 8 мг%, а в крови - кетоновые тела до 15 - 70 мг% и более при норме 1 - 6 мг%. Содержание сахара (глюкозы) в крови больных коров снижается до 30 мг% и ниже при норме 40 - 60 мг%, а резервная щелочность - ниже 40 об.% С02. В крови больных кетозом коров увеличивается уровень ЛЖК, НЭЖК, молочной, пировиноградной кислот, тироксина. В рубцовом содержимом отмечают сдвиг рН в кислую сторону, повышение концентрации масляной кислоты, аммиака, кетоновых тел.
При хроническом кетозе и вторичной остеодистрофии отмечают незначительное повышение содержания кетоновых тел в крови, снижение гемоглобина, мочевины, кальция, резервной щелочности, сахара, а также повышение общего белка, активности аспартатаминотрансферазы и лактатдегидрогеназы.
Билирубин - желчный пигмент, который образуется в клетках РЭС из гемоглобина разрушенных эритроцитов. В плазме крови он образует непрочный комплекс с альбумином, его называют свободным или непрямым. Уровень его в крови составляет 1-14 мкмоль/л. Из организма выводится через печень, в ней происходит экстрагирование билирубина из комплекса с альбумином и его соединение с глюкуроновой кислотой. Комплекс билирубина с глюкуроновой кислотой называют прямым билирубином. Он выделяется в желчь и поступает в кишечник, где превращается в уробилиноген. В сыворотке крови здоровых животных содержится в основном непрямой билирубин (до 80 % от общего). Уровень общего билирубина повышается при гемолитической желтухе, усиленном гемолизе эритроцитов, в меньшей степени - при гепатите и циррозе печени. Прямой билирубин находится в крови в незначительных количествах (от 0 до 5 мкмоль/л) и повышается при болезнях печени и ее выводящих путей.
Глюкоза (сахар) - основной источник энергии для многих клеток организма. На ее долю приходится более 90 % всех низкомолекулярных углеводов. Относительно постоянный уровень глюкозы в крови (40 - 60 мг% или 2,2 - 4,0 ммоль/л) поддерживается за счет действия гормонов. Гипогликемия встречается при кетозе, вторичной остеодистрофии, послеродовом парезе, некоторых формах ожирения, токсических поражениях печени. Часто она является следствием недостатка в кормах легкоусвояемых углеводов, большой потребности в глюкозе при высококонцентратном типе кормления, преобладания в рационе кислых кормов. Гипергликемия может быть стойкой (при сахарном диабете) и непродолжительной (при скармливании скоту больших количеств сахаристых кормов, а также при испуге, высокой температуре, стрессах).
Концентрация глюкозы в сыворотке (плазме) крови быстро снижается, поэтому ее определяют сразу после взятия крови или проводят осаждение белков трихлоруксусной кислотой непосредственно на ферме. На практике об этом часто забывают, что служит причиной неверных рекомендаций.
В лаборатории ЗАО "Витасоль" в 2004 -2006 гг. исследовали минеральный состав крови 4765 клинически здоровых коров из 12 регионов России. Установили, что содержание кальция варьировало в пределах 9,0 - 13,0 мг% и в среднем составляло 10,5 мг%, неорганического фосфора - соответственно 4,0 - 8,0 и 5,5 мг%, магния - 2,0 - 3,5 и 2,4 мг%, меди - 60,0 - 145,0 и 95,0 мкг%, железа - 110,0 - 220,0 и 170,0 мкг%, цинка - 65,0 - 160,0 и 105,0 мкг%.
Результаты исследований нашей лаборатории хорошо совпадают с данными литературы последних 10 лет (С.Г. Кузнецов, 1996; С.-х.. животные. Физиологические и биохимические параметры организма. Справочное пособие. Боровск, 2002), но отличаются от опубликованных в ранее изданных справочниках по ветеринарии (Клиническая лабораторная диагностика в ветеринарии. - М., 1985; Справочник по вете-ринарной биохимии. - Минск, 1988). Следует обратить особое внимание на повышение методического уровня работников ветеринарных лабораторий всех уровней и на периодический контроль их деятельности. Нередко в протоколах испытаний содержание кальция в крови клинически здоровых коров приводится ниже 3 мг% и специалистам хозяйств даются соответствующие рекомендации. Однако при повторном анализе крови от тех же животных в другой лаборатории получают совершение иные результаты.
Журнал "Ветеринария" №4 2008