

В.И. Глазко
д.с.-х.н., профессор, академик РАСХН Российский государственный аграрный университет - МСХА имени К.А. Тимирязева Россия, Москва
Аннотация
Рассматриваются собственные и литературные данные о результатах использования современных методов «селекции с помощью маркеров» (MAS) в животноводстве. Обсуждаются направления увеличения их эффективности, в частности, путем выявления геномных мишеней естественного отбора, усложняющих выявление геномных элементов, полиморфизм которых прямо ассоциирован с изменчивостью хозяйственно-ценных признаков. В этих целях предлагается использование для геномного сканирования полилокусное генотипирование наиболее полиморфных геномных элементов, фрагментов эндогенных ретровирусов.
Современные задачи ускоренной индустриализации животноводства, увеличения производства животноводческой продукции по отношению к затратам, требуют развития новых подходов к управлению генетическими ресурсами животных. В этой связи большие надежды возлагаются на современные достижения в области биотехнологий. Однако инновационные технологии в животноводстве только тогда становятся инновационными, когда приводят к увеличению прибыли на единицу затрат. В этом отношении широкое внедрение современных методов геномики и клеточных технологий в селекционную работу с сельскохозяйственными видами часто сопровождаются неоправданными расходами. Это, с одной стороны, сопровождается невосполнимыми экономическими потерями, с другой - приводит к утрате доверия практиков к внедрению современных молекулярно-генетических методов в животноводство.
Основные причины преувеличения возможностей экономического успеха применения современных.методов клеточной и молекулярной биологии в работе с животными сельскохозяйственных видов обусловлены недостаточной изученностью оптимальных Областей их использования и, соответственно, диапазона их эффектов. До сих пор остаются дискуссионными результаты внедрения таких методов, как селекция с помощью маркеров (Marker Assistant Selection - MAS), геномное сканирование и геномная селекция, множественная овуляция и трансплантация эмбрионов {multiple ovulation and embryo transfer - МОЕТ), сексирование семени у крупного рогатого скота [19].
Очевидно, что главная цель селекционной работы заключается в подборе таких пар для скрещиваний с высокой племенной ценностью, чтобы в следующем поколении добиться прогнозируемого селекционного успеха. До настоящего времени большинство программ по селекционному усовершенствованию, в частности, крупного рогатого скота, основывается на статистическом анализе больших баз данных по связям между генотипами и фенотипическими характеристиками у информационных модельных семей животных, на основании которых в дальнейшем делается прогноз развития желательных признаков у носителей определенных генотипов. Однако, долгая история исследований в области использования генетических маркеров для таких прогнозов свидетельствует об ограниченных возможностях их прямого внедрения в практические программы разведения. Появление методов геномного картирования высокоплотного распределения мононуклеотидных замен {Single Nucleotide Polymorphisms -SNP), объединенных с новыми статистическими обработками получаемых данных для ранних оценок племенной ценности животных, позволило сформулировать новое направление MAS, получившее название «геномной селекции». Предполагалось, что именно методы генотипирования по десяткам тысяч SNP позволят выявить геномные участки, полиморфизм которых ассоциирован с изменчивостью проявления желательных фенотипических признаков. Однако, в достаточно обширных исследованиях было показано, что генетическая обусловленность изменчивости признаков существенно варьирует от одного к другому, а также для большинства включаемых в анализ фенотипических признаков более 50% наследственной изменчивости попадают в геномные области с небольшими фенотипическими эффектами, порядок величин которых соответствует полигенному характеру наследования [12].
В наших собственных исследованиях выполнен анализ ассоциаций мононуклеотидных полиморфизмов (S/VP) в промоторах специализированных генов метаболизма липидов {Scd, Lep), миогенеза (Myod, Myf-6), гена регуляторного белка широкого спектра действия Орп) с показателями мясо-сальной продуктивности свиней [6]. Выявленные ассоциации зависели от происхождения животных (отличались у двух- и трех-породных кроссов) и их половой принадлежности. Наибольшее количество ассоциаций наблюдалось между SNP промотора гена Орп и толщиной шпика, характеристиками мясной продуктивности. Полученные данные свидетельствуют о том, что исследованные SNP генотипы действительно могут быть использованы для увеличения эффективности прогноза продуктивного потенциала животных, однако воспроизводимость таких ассоциаций ограничивается общностью происхождения, половой принадлежностью.
Сходные данные были получены нами при генотипировании пород крупного рогатого скота молочного и двойного направлений продуктивности по аллельным вариантам ряда генов (каппа казеин, бета-лактоглобулин, соматотропный гормон, фактор регуляции транскрипции - Pit-1, лептин), ассоциированных с характеристиками молочной продуктивности (общий удой, процент жира, процент белка и т.д.). Так, выявлена выраженная межпородная дифференциация по частотам встречаемости «молочных» аллелей; их частота статистически достоверно выше у молочных пород по сравнению с породами двойного направления продуктивности - однако у индивидуальных молочных животных не обнаруживается сцепления между присутствием желательных «молочных» аллелей по разным локусам [4].
Подобные результаты были представлены в работах французских исследователей по использованию множественного генотипирования по SNP для выявления аллельных вариантов, ассоциированных с. молочной продуктивностью у трех французских специализированных молочных пород [14]. Сопоставлено распределение SNP по хромосомным картам, неравновесие по сцеплению между ними, ассоциации гаплотипов с изменчивостью характеристик молочной продуктивности у голштинов, нормандов и монтбельярдов. Выделено 13 участков хромосом с наибольшей плотностью SNR аллельные варианты которых статистически достоверно ассоциированы с характеристиками молочной продуктивности, проанализированы локализованные в них гены. Оказалось, что гены, полиморфизм которых ассоциирован с характеристиками молочной продуктивности, отличаются у разных пород. Так же как и в наших исследованиях генотипов по промоторам генов контроля липидного обмена и миогенеза у свиней, такие ассоциации генотипов с характеристиками продуктивности наблюдались, но ограничивались спецификой происхождения животных [6].
Накопленные данные наглядно свидетельствуют о том, что в искусственный отбор по одним и тем же признакам продуктивности вовлекаются аллельные варианты разных генов, продукты которых участвуют в их формировании. Более того, по-видимому, имеются сложные межгенные взаимодействия, способствующие определенному антагонизму между аллелями разных генов, участвующих в желательном проявлении, в част; ности, таких характеристик, как общий удой. Этим могут объясняться межпородные отличия по частотам «мо-
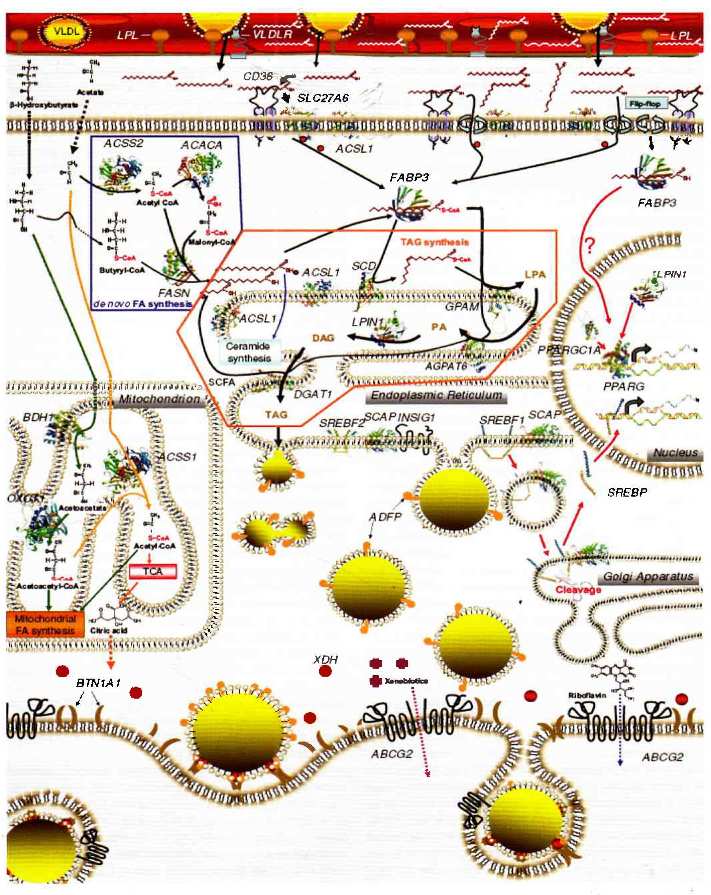
Рисунок 1. Схема взаимосвязей между клеточными путями синтеза и секреции, регулирующими синтез молочного жира в молочной железе крупного рогатого скота [8]
лочных» аллелей при отсутствии внутри породных ассоциаций между ними - внутри специализированных молочных пород не удается найти животных с комплексными генотипами, в которых объединены такие аллели по разным генам.
В пользу этого предположения свидетельствуют полученные нами данные об индивидуальной изменчивости органоспецифичных профилей генной экспрессии (ПГЭ) у свиней. Оказалось, что ПГЭ, в частности, печени, по индивидуальной изменчивости отчетливо разбивается на две компоненты - конститутивную и вариабельную. Обнаружено, что в среднем продукты генов вариабельной части ПГЭ одновременно участвуют в два раза в большем количестве метаболических путей по сравнению с конститутивной группой [5, 16]. Участие в большом количестве метаболических путей может приводить к индивидуальной изменчивости вклада продукта конкретного гена в формирование определенного фенотипического признака и, соответственно, к разным ассоциациям между генетически детерминированным полиморфизмом и фенотипической изменчивостью.
Таким образом, получается, что надежность ассоциаций между полиморфизмом определенных генов и желательным проявлением фенотипического признака зависит не только от биохимической функции их продуктов, но и сложности метаболических сетей, в которых они принимают участие. Из этого следует, что эффективность MAS прежде всего определяется положением генов и регуляторных генетических элементов в иерархии генных сетей, лежащих в основе формирования фенотипических признаков интереса. Особенности такой иерархии зависят от исследуемого признака. Большинство хозяйственно ценных признаков являются количественными, как правило, с низким коэффициентом наследования. В изменчивость их проявления основной вклад вносит вариабельность состава и направления действия факторов окружающей среды.
Наглядной демонстрацией сложностей в организации метаболических и генных сетей может служить описанные в литературе пути синтеза молочного жира в молоке крупного рогатого скота (рис. 1, 2) [8].
Следует отметить, что изменчивость такой характеристики молочной продуктивности, как процент жира, имеет существенно большую генетическую компоненту по сравнению с общим удоем (судя по общепринятым коэффициентам наследуемости -0,60 для процента жира по сравнению с -0,18 для общего удоя). То есть, сложность генных сетей, участвующих в контроле такой характеристики как общий удой, будет существенно больше, чем сетей, контролирующих синтез молочного жира (рис. 1, 2).
Именно этим, по-видимому, объясняется достаточно низкая эффективность многолетних попыток выявить главные гены молочной продуктивности, которая рассматривается как сочетание, в основном, величин общего удоя, процента белка, процента жира. Меняются методы геномного сканирования (полилокусного генотипирования), способы обработки получаемых данных и оценок ассоциаций между аллельными вариантами, а универсальных генотипов, позволяющих существенно улучшить оценки племенной ценности животных и его прогноза у молодняка не удалось найти до сих пор. Так, в последние годы широкое распространение получил новый метод геномного сканирования - оценка изменчивости по числу копий коротких геномных участков (Copy Number Variability, CNV - делеции, дупликации, транслокации, инверсии длиной от 100 до 1000 п. о.). Поскольку известно, что в мононуклеотидных заменах (SNP) участвует существенно меньшее количество нуклеотидов (~1/на 50000 пар оснований, п.о.) чем в изменчивости по CNV (~1/ на 10000 п. о.), с целью выявления и картирования главных генов молочной продуктивности выполняются геномные сканирования по распределению CNV вдоль хромосом у быков голштинской породы [21]. Д. Веллер, М. Рон - израильские исследователи, которые в начале 90-х годов первые пытались картировать главные гены молочной продуктивности с помощью микросателлитов на хромосомах крупного рогатого скота, уже обсуждают эффективность прямого включения данных по геномному сканированию в селекционные программы для отбора бычков без их оценки по потомству для сокращения среднего интервала между поколениями только по генотипированию критических для главных генов молочной продуктивности SNP (actual quantitative trait nucleotides -QTN). [25]. Предлагается также одновременно использовать разные методы полилокусного генотипирования (геномного сканирования), не только по SNP, сравнительной геномной гибридизации (comparative genomic hybridization - CGH), но и по CNV, поскольку каждый из этих методов позволяет получать дополнительные к другим данные об особенностях полиморфзима различных геномных элементов [26]. Тем не менее, эти новые подходы, связанные с увеличением «разрешаемости» методов ДНК секвенирования - точности выявления не только мононуклеотидных замен, но и изменчивости по копийности коротких геномных участков, не приводят к качественному изменению эффективности MAS.
Можно ожидать, что одной из причин, препятствующих успешному внедрению методов MAS в селекционную практику, является тот факт, что в реализацию желательного развития признака продуктивности вовлекается множество генов, продукты которых участвуют в контроле иммунной системы, внутриклеточного энергетического метаболизма, клеточных сигнальных путей, как это наблюдается при анализе изменений профилей генной экспрессии в процессе лактации у крупного рогатого скота [7]. Очевидно, что большое ког личество таких генов соответствует сложности изменений архитектоники молочной железы при лактации, а также питательной и защитной функций самого молока, возникших в процессе эволюции [17].
Интересно отметить, что геномные сравнения между яками и европейскими породами крупного рогатого скота позволили выявить отличия в генных семействах, продукты которых связаны не только с окислительным стрессом, но и с теми же клеточными сигнальными системами, ферментами внутриклеточного энергетического метаболизма, которые связывают с отличиями между этими видами по адаптированности к высокогорной гипоксии [20].
Накопленные данные позволяют сделать вывод о том, что в большинстве своем системы, существенные для процесса лактации у млекопитающих, являются частью систем общего метаболизма и иммунитета, которые участвуют в адаптации животных к меняющимся условиям окружающей среды. Такое заключение соответствует данным об изменчивости оценок племенной ценности быков по молочной продуктивности их дочерей в зависимости от места рождения дочерей, о сложных взаимоотношениях между устойчивостью к экологическим условиям разведения и продуктивностью животных мясных пород крупного рогатого скота [9]. То есть, геномные мишени естественного и искусственного отборов существенно отличаются между собой. Причем, кажется очевидным, что количество генов, продукты которых прямо участвуют в контроле, например, качества молока, такие как сывороточные и мицеллярные белки молока, может быть существенно меньше, чем вовлеченных в системы общего энергетического метаболизма и функций иммунной системы, связанных с адаптацией к меняющимся условиям окружающей среды. В наших собственных исследованиях, например, было обнаружено, что группы крупного рогатого скота одной и той же породы, воспроизводящиеся в разных эколого-географических условиях, не отличаются по частотам встречаемости аллелей ряда белков молока, несмотря на выраженные отличия по распределению аллелей генов, связанных с адаптивным потенциалом животных (рецептор витамина Д, пурин нуклеозидфосфорилаза) [4].
По-видимому, основным препятствием к практическому использованию MAS является тот факт, что в общем случае желательное проявление признака продуктивности зависит, в большей степени, от особенностей сочетаний генотипов по тем генетическим элементам, которые являются мишенями естественного отбора. Это и может обуславливать относительно низкую эффективность использования геномных участков с высокой плотностью SNR ассоциированных с племенной ценностью у быков, оцененных по потомству, для прогноза племенной ценности молодых животных, несущих те же генотипы, но сперма которых используется для оплодотворения в других эколого-географических регионах. Из этого следует, что для увеличение эффективности MAS необходимо накапливать информацию о геномных мишенях естественного отбора у животных сельскохозяйственных видов, на геномном фоне которых генетические элементы, прямо связанные с искусственным отбором, могут иметь разное фенотипическое проявление.
В этих целях необходим поиск геномных элементов, полиморфизм которых наиболее высок и в большей степени по сравнению с другими может вносить вклад в фенотипическую изменчивость. В этом отношении особый интерес представляет оценка геномного полиморфизма по диспергированным повторам, поскольку в настоящее время предполагается, что именно полиморфизм по этим генетическим элементам является наиболее широко распространенным вариантом генетической изменчивости. Так, известно, что большая часть генетического материала, в частности, у животных, представлена ретротранспозонами, в секвенированных геномах крупного рогатого скота они занимают 46,5% [22], домашней лошади - 36% [24]. Выраженный полиморфизм и видоспецифичность ретротранспозонов, высокая скорость их дивергенции даже за короткое время расхождения групп организмов от общего предка, наглядно описанная на примере геномов лабораторных линий мышей [18], позволяют полагать, что основным источником геномной изменчивости являются мобильные генетические элементы.
Следует отметить, что одной из выраженных особенностей геномного размножения ретротранспозонов является волнообразная динамика их возникновения, распространения и деградации. Для целого ряда ретротранспозонов описаны эволюционные циклы, включающие вертикальную или горизонтальную передачу, вспышку транспозиций с последующим разрушением большинства первоначальных копий [10]. Следами таких циклов является присутствие в хромосомах разных видов их множественных остатков [2,15, 23].
Мобильные генетические элементы могут перемещаться по геному с высокой частотой (по сравнению с источниками других мутаций), со скоростью в пределах от 10-3 до 10-5 на один элемент на одно поколение (даже до 10-2 при некоторых специфических скрещиваниях). Тем не менее, судя по полученным нами данным, а также имеющимся в литературе, сохраняется определенное постоянство близкой локализации, повышенной частоты рекомбинаций между разными эндогенными ретровирусами. Полученные данные открывают новые возможности для геномного сканирования путем использования в качестве геномных «якорей» участков длинных терминальных повторов эндогенных ретро в и русо в.
Ранее нами были получены данные, свидетельствующие о том, что одним из источников высокого полиморфизма фрагментов ДНК, фланкированных инвертированными повторами микросателлитов (ISSR-PCR маркеры) может быть их формирование
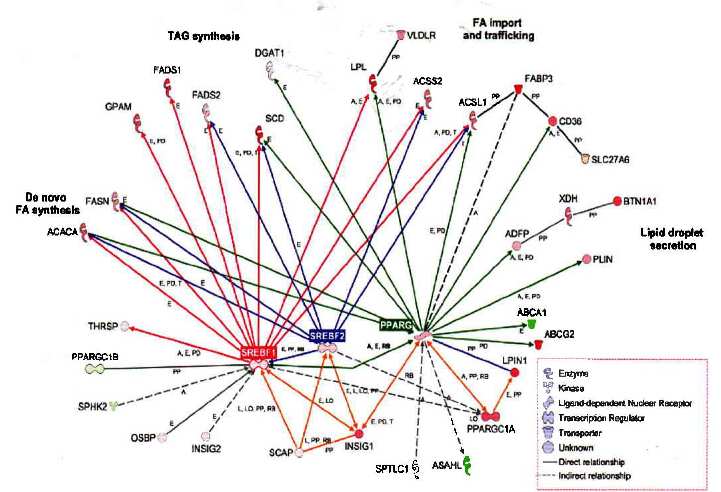
Рисунок 2. Сетевые взаимоотношения между генами, участвующими в синтезе молочного жира (http://www.ingenuity.com). Красные кружки отмечают положительные изменения, зеленые - негативные в экспрессии на 60 день лактации относительно -15 дня. Красные, голубые и зеленые стрелки отмечают гены, транскрипция которых находится под контролем регуляторных факторов SREBF1, SREBF2, и PPARG, соответственно. Светло оранжевые стрелки объединяют сеть, включающую PPARG, PPARGC1A, LPIN1, INSIG1, и SCAP, которая контролирует экспрессию/функцию SREBF белков. Буквы между стрелками отмечают: действия на активность (А), экспрессию (Е), локализацию (L0), протеолиз (L), РНК связывание (RB), белок-ДНК связывание (PD), и белок-белок связывание (РР). Гены сгруппированы по их исходным функциям в синтезе молочного жира.
в результате рекомбинационных событий между мобильными генетическими элементами [1, 2]. Следует отметить, что в секвенированных последовательностях геномов крупного рогатого скота и лошадей мобильные генетические элементы, в частности, с длинными концевыми повторами (LTR), несущими последовательности промоторов, сайты инициации транскрипции, энхансеры, встречаются на порядок чаще, чем микросателлитные локусы [13, 24] и их полиморфизм является видоспецифичным [15]. Более того, в нашиххюбственных исследованиях было обнаружено, что в высоко полиморфных участках ДНК, фланкируемых ивертированными повторами микросателлитов и состоящих из последовательностей, гомологичных рекомбинантам между разными длинными концевыми повторами эндогенных ретровирусов, наблюдается относительно повышенная частота встречаемости нуклеотидных последовательностей, способных формировать неканонические структуры ДНК, в частности, квадруплексы, типичные для точек рекомбинации [11]. Причем, наблюдается смещение их локализации к флангам участков гомологии к эндогенным ретровирусам (местам рекомбинаций между ними), для которых характерно наличие нуклеотидных последовательностей, предрасположенных к формированию и шпилечных структур, типичных для нуклеодидных последовательностей, участвующих в процессах регуляции генной экспрессии [3]. В этой связи геномное сканирование с использованием оценок полиморфизм участков ДНК, в которых локализованы фрагменты ретротранспозонов с длинными концевыми повторами, являются наиболее перспективным методом в поисках мишеней естественного и искусственного отборов у животных сельскохозяйственных видов, поскольку маркируют наиболее полиморфные геномные элементы потенциально участвующие в регуляции транскрипции.
Литература
1. Глазко В.И., Гладырь Е.А., Феофилов А.В, Бардуков Н.В., Глазко Т.Т. ISSR-PCR маркеры и мобильные генетические элементы в геномах сельскохозяйственных видов млекопитающих// Сельскохозяйственная биология - 2013. - №2. - С. 71-76;
2. Глазко В.И., Феофилов А.В., Бардуков Н.В., Глазко Т.Т. Видоспецифичные ISSR-PCR маркеры и пути их формирования// Известия ТСХА - 2012. - №1. - С. 118 - 125;
3. Зыбайлов Б. Л., Глазко В.И. Геномная нестабильность и неканонические структуры ДНК//Известия ТСХА. - 2012. - выпуск 5. - С. 108 - 122;
4. Харченко П.Н., Глазко В.И. ДНК технологии в развитии агробиологии. М.: Воскресенье, 2006. 473 с;
5. Хлопова Н.С., Глазко ТТ., Глазко В.И. Конститутивная и вариабельная компоненты профилей генной экспрессии печени свиней// Вавиловский журнал генетики и селекции, 2011, Том 15, № 1, С.130 - 135;
6. Хлопова Н.С., Стефанон Б., Гуатти Д., Глазко ТТ., Глазко В.И. Мононуклеотидный полиморфизм генов-кандидатов контроля показателей продуктивности свиней// Доклады РАСХН - 2012. - №4. - С. 39 - 45;
7. Bionaz M, Periasamy К, Rodriguez-ZasSL, Hurley WL, Loor JJ A Novel Dynamic Impact Approach (DIA) for Functional Analysis of Time-Course Omics Studies: Validation Using the Bovine Mammary Transcriptome//PLoS ONE - 2012. -Vol.7, N.3-e32455;
8. Bionaz M., Loor J. J. Gene networks driving bovine milk fat synthesis during the lactation cycle// BMC Genomics - 2008. - Vol. 9. - P. 366 - 380;
9. Burrow H. M. Importance of adaptation and genotype x environment interactions in tropical beef breeding systems//Animal. - 2012. - Vol. 6, N.5. - P. 729-740;
10. Carareto C.M.A. Tropical Africa as a cradle for horizontal transfers of transposable elements between species of the genera Drosophila and Zaprionus// Mobile Genetic Elements - 2011. - Vol.1, N.3. - P. 179-186;
11. Collie G.W., Parkinson G.N. The application of DNA and RNA G-quadruplexes to therapeutic medicines. //Chem. Soc. Rev. - 2011. - Vol.40. - P. 5867-5892;
12. Dekkers J.C.M. Application of Genomics Tools to Animal Breeding//Current Genomics. - 2012. - Vol. 13. - С 207-212;
13. Elsik C.G., R.L. Tellam, K.C. Worley. The Genome Sequence of Taurine Cattle: A Window to Ruminant Biology and Evolution // Science. - 2009. - Vol. 324, N. 5926. - P. 522-528;
14. Flori L, Fritz S., Jaffrezic F., Boussaha M. et al. The Genome Response to Artificial Selection: A Case Study in Dairy Cattle // PLoS ONE, 2009. Vol. 4. №.8. e6595;
15. Garcia-Etxebarria K., Jugo B.M. Genome-Wide Detection and Characterization of Endogenous Retroviruses in Bos taurus_// J- Virol. - 2010. - Vol. 84. - P. 10852- 10862;
16. Khlopova N.S., Glazko T.T Individual differences in gene expression in liver and kidney (Sus scrofa) In: Proc. of the International Moscow conference on computational and molecular biology (MCCMB'll), July 21-24,2011, Moscow, Russia - 2011. - P. 163 - 165;
17. Lemay D.G., Rijnkels M., German J.B.Lessons from the Bovine Genome: Implications for Human Nutrition and Research// J Nutr. - 2009. - Vol.139, N.7. - P. 1271-1272;
18. Nelleker C, Keane T.M., Yalcin B. et al. The genomic landscape shaped by selection on transposable elements across 18 mouse strains// Genome Biology - 2012 - Vol.13., R45. - P. 1- 21;
19. Pedersen L.D., Kargo M., Berg P., Voergaard J., Buch L.H., Siurensen A.C. Genomic selection strategies in dairy cattle breeding programmes: Sexed semen cannot replace multiple ovulation and embryo transfer as superior reproductive technology//! Anim. Breed. Genet. - 2012. - Vol.129 - P. 152-163;
20. Qiu Q., Guojie Zhang G., Ma T. et al. The yak genome and adaptation to life at high altitude//Nature Genetics - 2012. - Vol. 44, N.8. - P. 946 - 951;
21. Seroussi E., Glick G., Shirak A., Yakobson E., Weller J.I., Ezra E., Zeron Y. Analysis of copy loss and gain variations in Holstein cattle autosomes using BeadChip SNPs. //BMC Genomics - 2010. - Vol.11. - P. 673 - 693;
22. Tellam R. L, Worley К. С The Genome Sequence of Taurine Cattle: A Window to Ruminant Biology and Evolution//Science. - 2009. - Vol. 324. - P. 522 - 528;
23. Van der Kuyl A.C. Characterization of a Full-Length Endogenous Beta-Retrovirus, EqERV-Betal, in the Genome of the Horse (Equuscaballus)//Viruses- 2011. -Vol.3. - P. 620-628;
24. Wade СМ., Giulotto E., Sigurdsson S. Et al. Genome sequence, comparative analysis, and population genetics of the domestic horse / / Science - 2009. - Vol. 326, N.5954. - P. 865-867;
25. Weller J., Ron M. Invited review: quantitative trait nucleotide determination in the era of genomic selection.// J. Dairy Sci. - 2011. - Vol.94, N.3. -1082-1090;
26. Zhan В., Fadista J., Thomsen В., Hedegaard J., Panitz F., Bendixen С Global assessment of genomic variation in cattle by genome resequencingand high-throughput.genotyping//BMC Genomics - 2011. - Vol.12 - 557 (http://www.biomedcentral.com/1471-2164/12/557).
журнал VetPharma №2 2013