
Беляков А. А., Федорова Н. Н., Каниева Н. А., Грушко М. П.
ФГБОУ ВО «Астраханский государственный технический университет»
Введение
Благодаря своим высоким пищевым качествам шемая давно привлекала внимание рыбоводов [1, 3-5]. В 1929 г. начал искусственное разведение шемаи рыбовод А. Ф. Ершов на четырех рыбоводных пунктах в бассейне Кубани - реках Псекупсе и Пшише. Уже в 1968-1969 годах было выпущено в Азовское море более 10 млн штук молоди с навеской до 0,1 г. К 1971 году акклиматизация шемаи в низовьях Кубани - и Сенгилеевском водохранилище прошла успешно: шемая стала нереститься в естественной среде [5].
На сегодняшний день в научной литературе сведений о формировании тканей и органов шемаи в эмбриональном развитии представлено достаточно мало [2].
В связи с вышесказанным, целью иссле-дования явился анализ формирования тканей и систем органов шемаи на 7-м этапе эмбрионального развития (44-48 часов после оплодотворения при Т=23-24°С).
Материалы и методы
Работа выполнена на кафедре гидробиологии и общей экологии Астраханского государственного технического университета в 2015-2016 годах. Основными методами исследования были: ихтиологические (морфологические исследования зародышей шемаи), гистологические (изготовление и анализ серий срезов зародышей шемаи 44-48 часов после оплодотворения), статистические (математическая обработка цифрового материала). Объектом исследования служили зародыши шемаи, возраст которых составлял 44-48 часов после оплодотворения, при выращивании при температуре 23-24°С в аппаратах П. С. Ющенко. Сделано 20 фронтальных и сагиттальных серий срезов по общепринятым гистологическим методам. Сбор материала был проведен на Темрюкском рыбоводном заводе (Краснодарский край). Длина зародышей - 0,5-0,7мм, масса - 0,8 мг.
Результаты и обсуждение
Формирование нервной системы на 7-м этапе эмбрионального развития
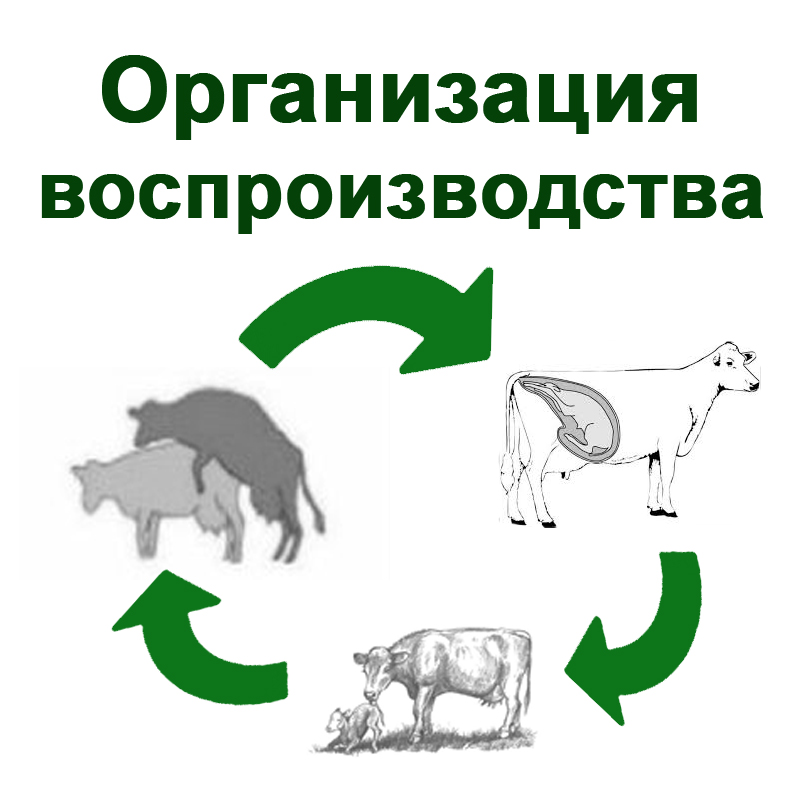
Зародыш был свернут в клубок вокруг широкой части желточного мешка (рис. 1). На стадии 44—48 часов после оплодотворения спинной мозг зародыша представлял собой полый тяж округлой формы, проходящий вдоль туловищного и хвостового отделов. Сегментарно вдоль спинного мозга располагались спинномозговые узлы (рис. 2).
У зародышей в возрасте 44-48 часов после оплодотворения серое вещество было представлено плотным скоплением нейробластов, белого вещества было очень мало - едва заметная полоска на периферии спинного мозга.
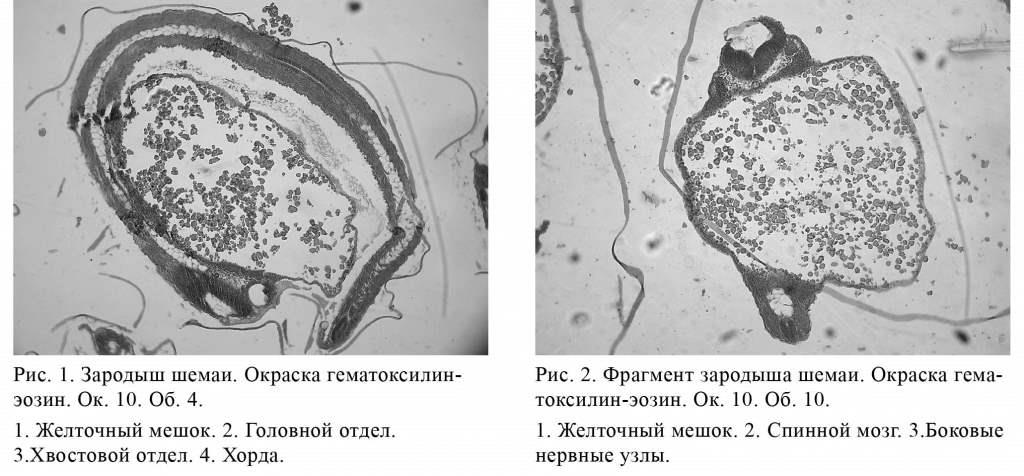
На изученных стадиях у зародышей на-метились все отделы мозга, но самым длин-ным оказался продолговатый мозг; из всех желудочков мозга наиболее широкой была ромбовидная ямка, особенно нижний ее треугольник. Полость третьего желудочка также напоминала треугольник, но была в три раза меньше ромбовидной ямки. Кроме того, имелась мезенхимальная закладка оболочек мозга и спинномозговых узлов.
Онапредставляласобойтканьснеболыпим количеством клеток и значительным количеством основного аморфного вещества. Полость спинного мозга у зародышей 44-48 часов была овальной в верхней части и округлой - в нижней, в 7 раз уже ромбовидной ямки. Хорда на этих стадиях развития была относительно широкой.
Органы чувств шемаи на 7-м этапе эмбрионального развития
У зародышей в возрасте 44-48 часов после оплодотворения четко выделить все три оболочки глазного яблока не удалось. Имелась общая закладка фиброзной и сосудистой оболочки из молодой соединительной ткани и плотная закладка сетчатки из 3-х рядов нейробластов. В глазном яблоке четко выделялась двухслойная эпителиальная роговица. У хрусталика имелась многоклеточная оболочка из 3-4 слоев округлых клеток с гомогенным центром. Полость глазного яблока, окружавшая хрусталик, была небольшой. Пигментных клеток в глазном яблоке не было (рис. 3).
У зародышей в возрасте 44-48 часов после оплодотворения появились небольшие полые закладки обонятельных ямок и слуховых пузырей.
Развитие пищеварительной системы в эмбриональном периоде (7-й этап)
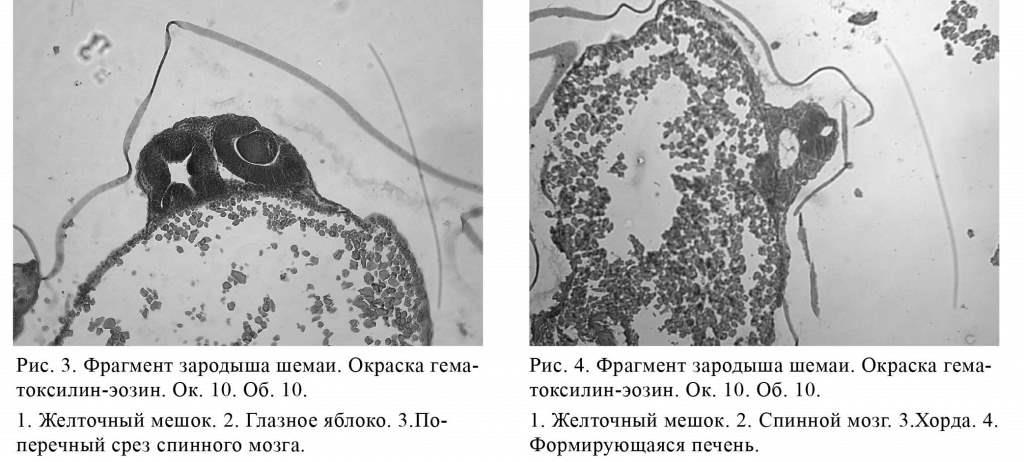
От уровня верхнего края желточного мешка позадней его стенке опускается зачаток будущего кишечника вдоль всей стенки мешка. Этот зачаток представляет собой трубку с очень незначительной полостью, равномерной на всем протяжении. Стенки зачатка покрыты высоким кубическим эпителием. Причем верхняя часть зачатка начиналась слепо, затем зачаток опускался по широкой части желточного мешка и заканчивался также слепо на уровне каудального конца узкой части желточного мешка. Под головным концом зародыша на верхнепередней части желточного мешка располагался плотный зачаток печеночной ткани (рис. 4). Формировавшаяся печень не имела трабекулярной архитектоники.
Большая часть желтка, находившегося в желточном мешке на 7-м этапе эмбриогенеза, оставалась неиспользованной. У желточного мешка имелись две оболочки: одна, верхняя, - соединительнотканная, под ней располагался кровеносный сосуд; нижняя, или внутренняя, - эпителиальная, из призматических эпителиальных клеток. В полости желточного мешка находились довольно крупные гранулы желтка, более мелкие из них плотно прилегали к эпителиальной оболочке желточного мешка.
Формирование сердечно-сосудистой си-стемы
У зародыша в возрасте 44-48 часов после оплодотворения было обнаружены в сердце две главные камеры - предсердие и желудочек. В брюшной и спинной аорте находились эритробласты.
Развитие мочеполовой системы Первичная почка у зародышей в возрасте 44-48 часов после оплодотворения представляла собой длинные парные образования, верхний уровень которых находился на середине желточного мешка, а нижний опускался до уровня каудального конца желточного мешка. Вдоль первичной почки опускался слепо начинавшийся мезонефридиальный проток. В строме первичной поч-ки находились сегментарно расположенные везикулы, основную массу стромы занимала кроветворная ткань.
Таким образом, как показали проведен-ные исследования, у зародышей в возрасте 44-48 часов после оплодотворения наиболее сформирован был продолговатый мозг в сравнении с другими отделами головного мозга. В этот же период у зародышей шемаи наиболее быстро формировался орган зре-ния, и происходило интенсивное развитие сердечно-сосудистой системы: главных ка-мер сердца - предсердия и желудочка.
Кроме того, в связи с формированием мезонефроса, в его межканальной ткани развивались клетки эритробластического ряда.
К 7-му этапу развития стала заметной плавниковая кайма, охватывавшая туловищный и хвостовой отделы зародыша.
Выводы
1. Развитие основных систем зародышей шемаи происходило гетерохромно. Причем на 7-м этапе эмбрионального развития была наиболее дифференцирована нервная система.
2. Из органов чувств у зародышей на 7-м этапе эмбрионального развития был сформирован орган зрения, хотя имелись закладки органов слуха и обоняния.
3. Мезонефрос на 7-м этапе эмбрионального развития функционировал как универсальный кроветворный орган.
Список литературы
1. Астанин, Л. П., Саманеева, Л. И. Сравнительное изучение плодовитости рыбца и шемаи, акклиматизированных в Сенгеевском водохранилище [Текст] / Л. П. Астанин, Л. И. Саманеева // Вопросы ихтиологии, 1968. - Т. 8. - № 4. - С. 646-653.
2. Беляков, А. А., Федорова, Н. Н., Грушко, М. П. Изменения клеток и тканей молоди шемаи (Chalcalbunus chalcoides) при искусственном вьфащивании [Текст] / А. А. Беляков, Н. Н. Федорова, М. П. Глушко // Труды ВНИРО : Аквакулыура, 2016. - Т. 162. - С. 139-145.
3. Битехтина, В. А., Карпенко, Г. И., Сафонова, М. В. Концепция воспроизводства шемаи в бассейне Азовского моря [Текст] / В. А. Битехтина, Г. И. Карпенко, М. В. Сафонова //1 конгресс ихтиологов России: тезисы докладов. -Астрахань : АГТУ, 1997. - С. 306.
4. Карпенко, Г. И. Экология шемаи Chalcalbunus chalcoides schischcovi Drensky в связи с ее искусственным разведением: дис. ... канд. биол. наук [Текст] / Г. И. Карпенко. -Москва, 1984. - 152 с.
5. Карпенко, Г. И., Швецова, Г. Н., Переверзева, Е. В., Головко, Г. В. Разведение шемаи в рыбоводных комплексах Азовского бассейна: технологическая инструкция [Текст] / Г. И. Карпенко, Г. Н. Швецова, Е. В. Переверзева, Г. В. Головко // Ростов-на-Дону : «Медиа-Полис», 2007. - 87с.