

Малашко В.В., Бапгура А.В.
УО «Гродненский государственный аграрный университет» г. Гродно. Республика Беларусь, 230008
Малашко Д.В.
«Белорусская государственная сельскохозяйственная академия» г. Горки, Могилевская обл.. Республика Беларусь. 213407
Введение. Лактационная функция присуща лишь женскому организму млекопитающих - одному из наиболее высокоорганизованных классов позвоночных. У всех видов млекопитающих длительность периодов одной беременности и одной лактации в сумме составляют, примерно, постоянную величину, колеблющуюся от 3,3% (у мелких видов) до 3,1% (у крупных) от общей продолжительности жизни [8]. Молочная железа и способность вскармливать новорожденных детенышей молоком является одним из довольно поздних филогенетических приобретений в эволюции позвоночных и, представляет пример важнейшего эволюционного скачка [2]. Формирование молочных же-лез в филогенезе, вероятно, шло по линии концентрации отдельных трубчатых желез в единый компактный трубчато-альвеолярный орган [3]. На последующих этапах филогенеза произошла дальнейшая специализация железистых органов, приспособившихся к выделению того или иного секрета. Развитие кожных желез можно представить в виде гипотетической схемы:
Неспециализированные кожные железы звероподобных рептилий
Потовые железы млекопитающих Сальные железы млекопитающих Молочные железы млекопитающих
Специфические железы
Закладка и развитие молочной железы у млекопитающих начинается уже на самых ранних этапах эмбриогенеза. Закладка молочной железы у эмбрионов всех млекопитающих, в том числе и человека, происходит по обеим сторонам живота в виде узких длинных полосок утолщенного эпителия, так называемых, млечных линий. К моменту рождения у большинства видов животных оказываются сформированными соски, связочный аппарат и междольковые перегородки. Основные же структуры паренхимы молочной железы, в том числе и протоки, носят еще рудиментарный характер. На месте будущей паренхимы находится жировая ткань, имеющая важное значение для процессов маммогенеза [5].
От рождения до половой зрелости развитие характеризуется прогрессивным ростом системы протоков. В этот период железа значительно увеличивается в размерах, но в основном это происходит за счет разрастания соединительной и жировой тканей. Наиболее интенсивный рост молочной железы начинается перед течкой.
Исследование морфологии секреторных клеток молочной железы на светооптическом и электронно-микроскопическом уровнях, показали, что молоко образуется в результате апокриновой, так и мерокриновой типов секреции. Голокриновый тип секреции - скорее исключение. При апокриновом типе секреции при выходе секрета из клетки апикальный участок последней вытягивается в просвет альвеолы и отделяется, высота эпителиальной клетки значительной уменьшается. Секрет выделяется в просвет альвеолы вместе с некоторыми другими компонентами клетки, а образовавшая его клетка вновь вырастает до прежней величины, после чего может начаться новый цикл секреции [9]. При мерокриновом типе секреции капля секрета (жира), продвигаясь к апикальной части клетки, оттесняет к базальной мембране цитоплазму и ядро. Апикальная плазмалемма обтекает жировую каплю и отрывается вместе с ней, не увлекая за собой фрагментов цитоплазмы. Мерокриновая секреция может сопровождаться выходом секрета через отверстия в клеточной оболочке или через неповрежденную оболочку (рис. 1).
Как отмечают И. И. Грачев и др. [4], на протяжении суток изменя-ется продолжительность латентного периода. Величина латентного периода у коров составляет 35-69 сек. [6]. Например, в утреннюю дой-ку молоковыделительная реакция наступает быстрее, чем в последую-щие часы. Это объясняется усилением сигнализации с рецепторов ор-гана в результате раздражения их в процессе накопления большого количества молока (за большее время между дойками) и повышения тем самым возбудимости центра молоковыведения.
Рефлекс выведения молока зависит от частоты и длительности раздражений, наносимых на рецепторы молочной железы. Оптималь-ной частотой механического раздражения рецепторов соска являются 90-130 сжатий соска в 1 мин. При такой частоте латентный период молоковыделительного рефлекса короткий и наблюдается наиболее полное проявление рефлекса: выделяется большое количество молока и скорость выведения значительно выше.
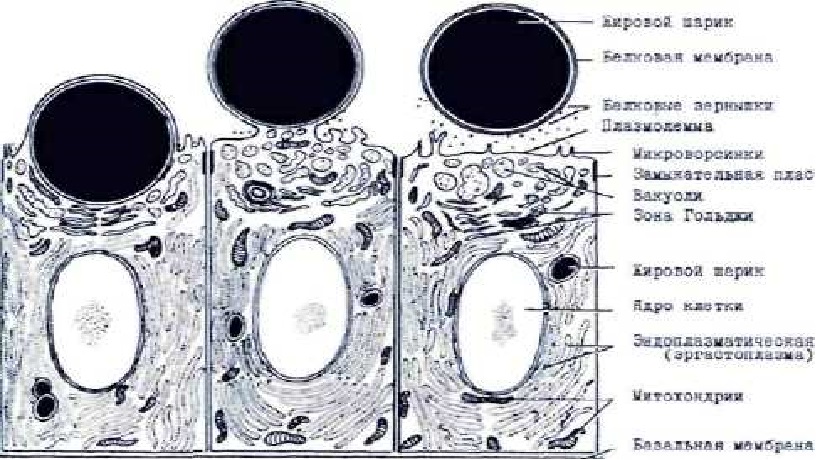
Рис. 1. Схема образования и выведения жировых шариков и белковых частиц в молочной железе (схема)
Процесс выведения молока при доении и сосании условно можно разделить на 2 фазы. В первую фазу выводится молоко, находящееся в цистерне и крупных выводных протоках (цистернальная порция). Эта фаза возникает через З-6 сек. от начала раздражения рецепторов молочной железы и продолжается первые 25-50 сек.
Во вторую фазу выводится молоко, находящееся в альвеолах и мелких выводных протоках. Вторая фаза является продолжением первой, она наступает через 25-50 сек. от начала раздражения рецепторов органа и может продолжаться 1-2 мин.
Всего на доение 1 коровы требуется затратить 8,5 мин., в т. ч. 1,5-2 мин. на подготовку, 6 мин. на доение и 30 сек. на заключительную об-работку. Если затрачивается больше времени на подготовку вымени, то, возможно, расстройство молокоотдачи. Например, в Голландии за 5 мин. корова отдает до 20 л молока, в Республике Беларусь - 10 л в среднем за 7 мин.
Для образования 100 мл молока через вымя должно пройти 54 л крови, азота в 100 мл молока надо через вымя пройти 45 л крови, фосфора в 100 мл молока через вымя проходит 47,6 л крови, лактозы в 100 мл молока через вымя протекает 25,6 л крови, жирных кислот в 100 мл через вымя должно пройти 48,8 л крови. В первую лактацию объем вымени составляет 17,33 дм3, а к 4 лактации - 33,70 дм3 и увеличивается на 94,5%, а в длину - на 22,5%, ширину - на 13,14% и глубину - на 51,4%. Масса вымени, примерно, составляет 15-25 кг, а без молока - 3% от массы тела.
Цель работы - исследование морфологических, физиологических особенностей и патологических процессов в молочной железы коров в процессе лактации.
Материал и методика исследований. Исследовались образцы молочной железы коров, всего было исследовано 47 тканевых образцов и 26 проб от клинически здоровых животных, выбракованных по при-чине болезней опорно-двигательного, пищеварительного и дыхательного аппаратов. Материал отбирался на ОАО «Гродненский мясокомбинат» на протяжении 2010-2013 годов. Для режущей прижизненной биопсии мягких тканей вымени использовали одноразовую стерильную иглу- COLT. Игла COLT позволяет взять размер столбика 20 мм.
Биоптаты вымени фиксировали в 10% нейтральном забуференным формалином по Р. Лилии, жидкости И. Карнуа, фиксаторе ФСУ А. М. Бродского, 70° спирте, а для проведения гистохимических исследова-ний биоматериал замораживали в жидком азоте (t-196°C) в сосуде Дьюара. Для получения обзорной информации структурных компо-нентов молочной железы гистосрезы окрашивали гематоксилин - эози-ном по П. Эрлиху, прочным зеленым по И. Ван Гизону, эозином - ме-тиленовым синим по Лейшману, альцнновым синим с докраской ядер гематоксилином. Микроциркуляторное русло молочной железы выявляли по методу В. В. Куприянова. Для электронно-микроскопического исследования ткань фиксировалась в 2% раствор глютарового альдегида. В последующем ткани помещали в 5% раствор глютарового альдегида на 2 часа. Глютаровый альдегид готовился на 0,1М фосфатном буфере рН 7,2-7,4 и фиксировали при t+4°C. Затем изготовляли кубики с длиной края 1-1,5 см. После 3-кратной промывки в ОДМ фосфатном буфере, материал обрабатывали 2%-ым раствором четырехокиси осмия, дегидрировали в спиртах, возрастающей концентрации, контрастировали уранил ацетатом и заключали в аралдит. Срезы готовили на ультрамикротоме ЛКБ (Швеция), контрастировали цитратом свинца и просматривали под микроскопом JEM -100В и JEM-100CX «JEOL» (Япония).
Результаты исследований и их обсуждение. Установлено, что 1 кг железистой ткани вымени за 15 часов вырабатывает 5-9 л молока, или за 1 час - 15 г жира, 15 г молочного сахара, 10 г белка, т. е. в сумме 40 г плотных веществ, примерно, 4% от массы железы. За 5 часов вымя синтезирует веществ равных собственной массе. Одним из недостатков вымени является неспособность противостоять инфекции в результате особенности строения: мало соединительной ткани, которая могла защитить от инфекции; отсутствует заметная фагоцитарная способность эндотелиоцитов капилляров, в отличие от печени, селезенки; выводная система вымени сообщается с внешней средой основным источником инфекции.
У коров в отличие от других животных в сосках обнаруживаются очень сложные и крупные чувствительные нервные окончания, очень похожие на окончания в руке человека. В соске различают основание, стенки и верхушку. Сосок содержит по одному протоку, у кобылы -два, у свиньи - 3, у собаки - 5-8 протоков. Проток открывается только во время доения или сосания, а цистерна по мере наполнения молоком. Емкостная система вымени (цистерны и протоковая часть) способна увеличиваться при заполнении молоком без повышения внутривыменного давления и в состоянии вместить в промежутках между дойками до 17-25 л молока. Радиально расположены складки, образующие цистернальную розетку, которые сохраняются лишь вокруг внутреннего отверстия соскового протока.
Сфинктер расположен на нижнем конце соска. Кончик соска очень чувствителен. Сосковая цистерна покрыта двурядным эпителием, ко-торый у внутреннего отверстия соскового канала резко переходит в многослойный ороговевающий плоский эпителий соскового канала. Сосковый канал известен еще под названием «полосатого канала». Для усиления сфинктера верхний конец соскового канала снабжен широкой складкой слизистой оболочки «розетка Фюрстенберга». Прижимаясь книзу и закрывая отверстие канала, складка препятствует вытеканию молока. У 19-37% коров такой складки не существует.
Сосковому сфинктеру отводится важная роль в двигательной функции молочной железы, например, расслабление соскового сфинк-тера, имеет определяющее значение для протекания первой фазы молоковыделительного процесса. От тонуса соскового сфинктера зависит скорость выведения молока. В особенностях строения соскового сфинктера кроется причина тугодойкости коров.
Причинами тугодойкости у коров могут быть перераздражение мышц сфинктера, чрезмерно развитая эластическая ткань в кончике соска или плотная соединительная ткань вокруг соскового протока, высокие складки в сосковой цистерне. Следует заметить, тугодойкость - наследственный порок. Например, фермеры США стремятся формировать стада не только из высокопродуктивных, но быстро и одновременно выдаиваемых коров. Скорость выдаивания - наследуемый при-знак. Однако это приводит к повышенной заболеваемости маститом. Важно отметить, что у тугодойких коров меньше соматических клеток в молоке.
В итоге нарушение процессов молоковыделения заключается в сужении канала соска, что ведет к тугодйкости или полному зарастанию (облитерации). При поражении соска обнаруживают: сосок становится меньше и более мягкий, чем здоровые, а соответствующая часть вымени больше, чем другие. Пальпацией обнаруживают сужения и закупорку сосковой цистерны в связи с утолщением слизистой оболочки или узелки (папилломы) и тяжи разросшейся соединительной ткани в канале соска (рис. 2а, б).
Степень растяжимости соскового протока колеблется в пределах 2,2-5,4 мм. Растяжимость соскового канала на 1 мм увеличивает минутный объем выдаиваемого молока, примерно, на 0,2 л и тем самым уменьшает длительность дойки на 2,2 мин. Диаметр соскового канала максимально может достигать 2,6-3,8 мм. Минимальное сопротивление соскового сфинктера составляет 30-143 мм рт. ст., утром оно больше, в обед и вечером меньше. Сильные возбуждения, такие как, охота, испуг, провоз корма и др. стресс-факторы тормозят расслабление сфинктера.
Продолжительность каждого сокращения сфинктера соска составляет 7-11 сек. (это составляет 6-9 колебаний в 1 мин.) и ритмические колебания сосковой цистерны с амплитудой 6-8 см вод. ст. и продолжительностью от 7 сек. до 12 сек. Сосковая цистерна вмещает 15-45 г молока. В передней доле вымени поверхность сосковой цистерны гладкая или имеет бухтообразные впячивания в сторону стенки соска.
Стенка соска от основания до сфинктера имеет одинаковую толщину. В задней доле вымени сосковая цистерна в области верхушки и в середине тела имеет гладкую поверхность. Ближе к основанию соска имеются бухтообразные впячивания в его стенку (рис. 3). Молочная цистерна имеет длину от 3 см до 8 см.
Скорость вытекания молока из соска при вакууме в линии 50 кПа -8,5 м/сек., при 40 кПа -7,5 м/сек. Физиологически эффективный диаметр открытого канала ~2 мм. Всасывающая сила молока, которая вызывает повреждение поверхности открытого канала соска -—1,8*10 ~2 N. Поверхности закрытого канала соска сразу после доения находятся на расстоянии в среднем 100 мкм друг от друга.
Поэтому бактерии, заражающие оставшееся в соске молоко, не могут находиться более чем на 50 мкм от поверхности его канала. В зад-ней части доли молочные ходы имеют дугообразное направление -назад, вверх и вперед, а в средней и передней частях - от молочной цистерны вверх. Исходя из такого расположения молочной цистерны задней доли, катетер должен вводиться по направлению снизу, вверх и назад, а не строго вертикально (рис. 4 и рис. 5). Во время доения температура на поверхности кожи вымени увеличивается на t+0,3-2,4°C, a внутри цистерны - на t+3-9°C, в тканях вымени - на t+O,25-O,85°C.
Кожа сосков вымени не имеет волос, потовых и сальных желез, но увеличивается содержание мышечной ткани и утолшение эпидермиса. Эпидермис кожи в соске имеет до 35-40 клеточных слоев (как результат механического воздействия на соски, например, для получения 1 л молока необходимо провести около 90-100-130 сжатий в мин.), но и врастает в дерму соска длинными отростками. Соски защищены от низкой температуры развитыми артерио - венозными анастомозами типа замыкающих артерий. Слизистая оболочка соска покрыта многослойным ороговевающим плоским эпителием, и слизистая оболочка становится менее прозрачной. Длина протока достигает 0,5-0,6-1,3-1,4 см.
Поверхность протока покрыта массой слущенных (десквамирован-ных) клеток. Они содействуют плотному закрытию просвета протока, и содержит бактериостатические и бактерицидные вещества, липиды, в первую очередь жирные кислоты (лавровую, миристиновую, олеиновую, линоленовую). Данный продукт имеет сходство с кожным салом, поэтому его называют «молочным салом - lactosebum». Лактосебум оказывает бактерицидное действие на Str. agalactiae и др. микробы. Повышенная кератинизация протока (особенно у старых коров) снижает защитные свойства лактосебума.
Барьерная функция сосков против микробов сразу после доения до-вольно значительная и медленно повышается в течение первых 30 мин. Через 120 мин. наблюдается падение защитной роли соскового канала (до минимальной) на 72-76%, после чего в интервале 120-240 мин. отмечается подъем барьерной функции, которая возрастает в период 4-8 час. после доения почти на 55% от первоначального периода. После дойки сосковый канал остается открытым в течение 1-2 часов. При повреждении сосков и при мастите они остаются частично открытыми на постоянной основе.
Повреждение кончика соска может происходить при повышенном давлении или колебании вакуума. Антибактериальные свойства соскового канала могут нарушаться при введении канюли шприца для инфузии лекарств. Дезраствор уничтожает бактерии на кончике соска, но часть микробов заносится в сосковую цистерну с канюлей шприца. Дезинфекция кончика соска не влияет на бактерии, заселяющие кера-тин соскового канала. При полном введении канюли шприца в соско-вую цистерну могут проталкиваться кусочки кератина с микробами, что может привести к возникновению инфекции. Обычно канюля шприца в 8 раз больше диаметра соскового канала, тем самым при его введении происходит растягивание канала, что способствует снижению устойчивости кератина к инфекции. Микробы в вымени локали-зуются преимущественно в цистернах и более крупных молочных про-токах, т. к. мелкие молочные протоки секретируют лизоцим, что пре-пятствует проникновению микробов в верхние части вымени. В мо-лочных цистернах и крупных молочных протоках температура более низкая, что создает «комфорт» для развития микробов.
Согласно физиологической норме сосковый канал должен оставаться открытым после машинного доения в течение 30^4-0 мин. По отдельным фенотипическим признакам сосков можно судить о резистентности коров к маститам. У коров с дискообразными концами сосков восприимчивость к маститу выше, чем у коров с овально закругленными концами. Если в содержимом канала сосков жирных кислот (лавровой, миристиновой) до 20 мг, то маститом коровы боле-ют редко, а преобладание в сосках олеиновой и линоленовой свидетельствует о высокой восприимчивости коров к маститу.
Существует прямо пропорциональная зависимость между частотой маститов и толщиной сосков, а также скоростью доения. Зависимость возникновения мастита и расстоянием от верхушки сосков до земли
носит обратно пропорциональный характер. Сосковые каналы, которые шире и короче служат одной из причин, повышенной заболевае-мости коров маститом.
На продуктивность животных и восприимчивость к маститу оказывает влияние форма сосков, примерно, цилиндрическую форму имеют 66,32%, конусообразная - 33,68% животных. Форма кончика соска может быть круглая, плоская, воронкообразная. Четверти вымени с цилиндрическими сосками имеют 59,3% коров, с круглыми кончиками - 35,1%, с плоскими и воронкообразными кончиками - 5,6%. Четверти вымени с конусообразными сосками продуцировали молоко, свободное от бактерий и содержащее соматических клеток менее 500 тыс/мл, чем с цилиндрическими сосками (68,2% и 54,8% соответственно), а пробы с наличием бактерий (18,9% и 20,4% соответственно), более 500 тыс/мл соматических клеток (12,9% и 24,8% соответственно). Соотношение тканей вымени коров, представлено в таблице 1.
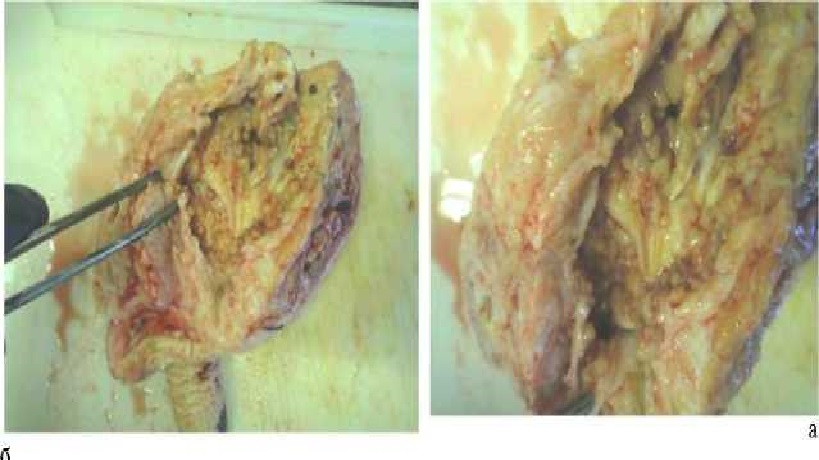
Рис. 2. Папилломы в молочной цистерне и сосковом канале коровы (фото оригинал А. В. Башуры).
440
Таблица 1. Соотношение тканей вымени коровы
|
Ткань |
Большое вымя, свыше 20 кг |
Среднее вымя |
Маленькое вымя, меньше 10 кг |
|
Интерстициальная ткань |
12.9 |
13.2 |
13,0 |
|
Жировая ткань |
7.9 |
8.1 |
8.2 |
|
Просветы альвеол |
62.7 |
59,7 |
61,2 |
|
Межальвеолярная ткань |
6,5 |
7,9 |
6.9 |
|
Итого: |
79,2 |
78,7 |
78,8 |
|
Паренхима железы: -количество желез на 3 мм2 -величина альвеол, мкм3 |
152 14934 |
122 16669 |
143 16041 |
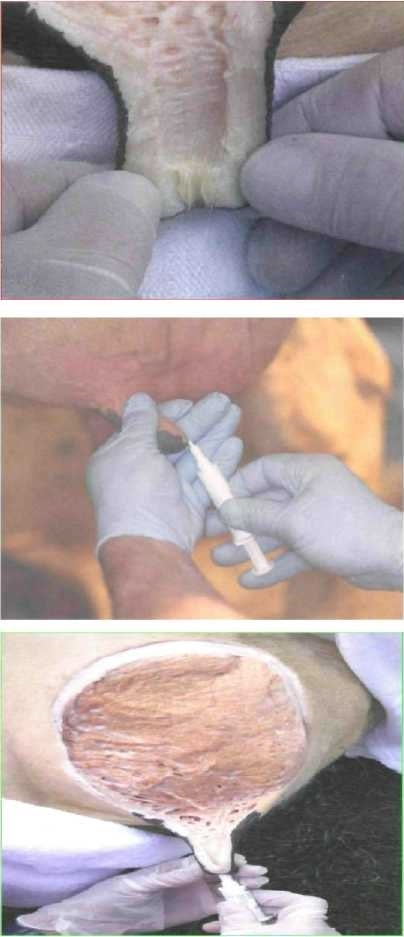
Рис. 3. Внутреннее строение соска коровы (продольны разрез)
Рис.4. Введение канюли шприца в сосковый канал.
Рис. 5. Введение канюли в сосковый канал (разрез вымени).
У старых коров количество интерстициальной ткани увеличивается до 23%, а жировой ткани уменьшается - с 11,4% до 8%. Важной клеточной структурой молочной железы являются миоэпителиальные клетки. Задача клеток - опорожнение альвеол и мелких протоков от молока.
Сокращение клеток начинается через 30-50 сек. после раздражения сосков. Функция клеток регулируется окситоцином (гормон задней доли гипофиза). Действие окситоцина продолжается в течение 5-7 мин., затем окситоцин окисляется и его влияние на клетки прекращается. Рефлекс, вызывающий выделение окситоцина может повториться у коров, не раньше, чем через 30 мин. после предыдущего рефлекса. Окситоцин практически не оказывает заметного действия на сократи-тельную способность миоэпителия в предродовый период формирования лактационной функции. На послеродовом этапе развития лактации роль окситошша сводится к эффективной и быстрой стимуляции функции миоэпителия альвеол вымени.
Важную роль молочная железа играет в иммунологическом процессе. В клетках эпителия молочной железы повышено число рецепторов к IgG. что способствует переходу IgG из крови в молозиво и при-соединению IgG к клеткам кишечника у телят. В процессе беременно-сти и после родов молочная железа имеет большое иммунологическое значение, поскольку в ней происходит как переход антител из сыво-ротки крови в молозиво и их концентрирование в ней, так и синтез различных классов иммуноглобулинов.
В состав молозива входят IgG, IgM, IgA, компоненты комплемента, лактоферрин. лизоцим, полиморфоядерные лейкоциты, моноциты, макрофаги, Т- и В- лимфоциты, плазматические клетки. IgA секрети-руется преимущественно плазматическими клетками молочной железы, IgM - частично местно, частично поступают из крови, IgG - почти целиком поступает из крови. Однако концентрация IgG в молозиве значительно превосходит его содержание в сыворотке крови [7].
Первичным сигналом для начала процесса роста молочных протоков служат экстрогены в комплексе с соматотропином гипофиза и стероидами надпочечников. Причем воздействие эстрогенов должно быть относительно длительным (от нескольких недель до нескольких месяцев), что, вероятно, соответствует периоду от начала полового созрвания самок до иx физиологической зрелости.
Для завершения роста протоков, альвеол и дифференцпровки секреторных клеток на определенном этапе необходимо подключение к вышеуказанным гормонам прогестерона и пролактпна. Прогестерон при наступлении полового созревания синтезируется в желтом теле, образующемся периодически, после овуляции яйцеклетки. Рост и развитие секреторного аппарата молочной железы завершаются во время беременности, когда наиболее интенсивно функционирует кора надпо-чечников, а соматотропин и пролактин продуцируются не только в гипофизе, но и в плаценте. Прогестерон во время беременности образуется в постоянно функционирующем желтом теле и плаценте.
В процессе массажа молочной железы, сосания или механического выведения молока сигналы по афферентным путям спинного мозга передаются в гипоталамус, затем в гипофиз, где в ответ выделяется окситоцин, вызывающий сокращения миоэпителиальных клеток аль-веол и протоков, а также (в начале кормления теленка или выведения молока) происходит быстрое массивное выделение пролактина, необ-ходимого для поддержания лактогенеза и накопления молока для следующего кормления.
Заключение. Следует учитывать, что для нормального развития и функционирования молочной железы, как у первородящих, так и у сухостойных животных необходимо последовательное воздействие на нее различных гормонов, участвующих в регуляции процессов роста протоков, альвеол, секреторных клеток молочной железы и секреции составных частей молока, в том числе специфических и неспецифических факторов резистентности.
ЛИТЕРАТУРА
1. Вальдман, Э. К. Моторная функция выменн коров при машинном доении / В. К. Вальдман//Докл. Советских ученых к ХГХ Всемир. конф. -М., 1971. -С. 71.
2. Галанцев. В. П. Эволюция лактации / В. П. Галанцев. Е. П. Гуляева. - Л.: Наука, 1987.-176 с.
3. Грачев. И. И. Физиология лактации сельскохозяйственных животных / И. И. Грачев, В. П. Галанцев. - М.: Колос, 1974. - 279 с.
4. Грачев, И. И. Цитофизиология секреции молока / И. И. Грачев. С. М. Попов, В. Г. Скопичев. -Л.: Наука, 1976. -238 с.
5. Зеликовская, 3. 3. О секреторном процессе в молочной железе/3. 3. Зеликовская. А. А. Туревский // Арх. анатомии, гистологии и эмбриологии. -1962. -№ 63. - С. 76-80.
6. Кавешникова. К. И. Физиологические основы машинного доения / К. И. Кавешникова//Вопросы физиологии машинного доения. -М.. 1970. -с. 25-32.
7. Петров, А. М. Формирование колострального иммунитета у животных / А. М. Петров //Ветеринария. -2006. -№8. -С. 35-41.
8. Blaxter. К. L. The comparative biology of lactation / К. L. Blaxter // Lactation. Nottingham. 1970. -P. 51-69.
9. Kurosumi. K. The fine structure of mammary glands of lactating rats, with special
reference to the apocrine secretion / K. Kurosiiini, N. Baba // Exp. Cell. Res. - 1986. -Vol. 50.
-P. 177-192.