
Н. И. КРИВЦОВ, Р. Б. КОЗИН, В. И. ЛЕБЕДЕВ, В. И. МАСЛЕННИКОВА. ПЧЕЛОВОДСТВО
Продукты расщепления питательных веществ, поступивших в клетки тела,могут передать содержащуюся в них энергию только при окислении. Необходимый для этого процесса кислород доставляется органами дыхания, которые одновременно удаляют из организма конечные продукты распада — диоксид углерода и воду в виде пара.
СТРОЕНИЕ ДЫХАТЕЛЬНОЙ СИСТЕМЫ
Дыхательная система насекомых своеобразна. Она состоит из большого числа сильно р азветвленных воздухоносных трубок — трахей, пронизывающих все тело. Трахеи открываются наружу с помощью дыхалец (стигм). Мельчайшие разветвления трахей образуют трахейные капилляры— трахеолы. У медоносных пчел крупные трахейные стволы образуют сильные расширения — воздушные мешки (рис. 23).
Трахея представляет собой трубку, стенки которой состоят из двух слоев: внутреннего хитинового (интимы) и внешнего однослойного трахейного эпителия (матрикса).
Внутренний хитиновый слой содержит тонкую экзокутикулу, которая образует спиральные утолщения (тенидии, или с пираль). Эти утолщения постоянно поддерживают трахею в расправленном состоянии. Одновременно спиральные утолщения придают трахеям прочность, гибкость и растяжимость. Так, при повышении давления трахея может укоротиться на 20-30% , а при растяжении — увеличиться почти вдвое без нарушения ее строения и функции.
Дыхальца, или стигмы, расположены по бокам сегментов груди и брюшка. Упчелы 10 пар дыхалец — три на грудном отделе и семь на брюшном.
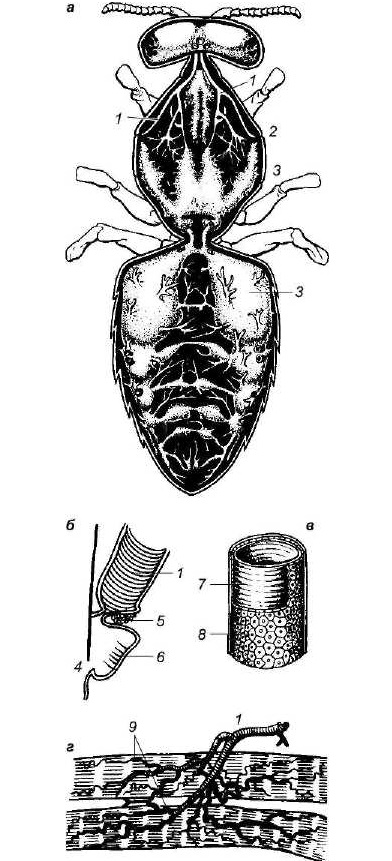
Рис. 23
Дыхательная система пчелы:
а — воздушные мешки и главные трахейные стволы; б— продольный разрез брюшного дыхальца; в — часть трахеи при сильном увеличении; г — разветвления трахеи и трахеолы в мышечной ткани; 1 — трахеи; 2 — первое грудное дыхальце; 3 — воздушные мешки; 4 — вход в дыхательную камеру; 5 — мышца замыкающего аппарата; 6 — волоски; 7 — спиральные утолщения; 8 — эпителиальные клетки; 9 — трахеолы.
С наружным воздухом дыхальце сообщается через небольшое продольное отверстие. Через это отверстие воздух попадает в сферическую полость — дыхательную камеру, внутренние стенки которой обильно покрыты волосками. Сеть волосков выполняет функцию фильтра, проходя через который, воздух очищается от механических примесей (зерен цветочной пыльцы).
Д. Ли (1963) показал, что молодые пчелы имеют в дыхальцах мягкие эластичные волоски, через которые легко проникает в трахею клещ акарапис вуди. У пчел старше 9-дневного возраста волоски затвердевают, и клещи через такую решетку волосков проникнуть не могут. Поэтому в этом возрасте пчелы не заражаются.
От переднего края дыхательной камеры в косом направлении отходшветвъ трахеи, которая открывается вторым концом в воздушном мешке. Отверстие трахеи имеет вид щели. У входа трахеи в дыхательную камеру расположен замыкающий аппарат, который состоит из двух пружинных кусков хитина (замыкающих конусов), связанных замыкающей мышцей. Сближаясь, края конусов закрывают трахею, и воздух перестает поступать в нее. К нижнему (хорошо развитому) конусу прикреплена еще размыкательная мышца, при сокращении которой вершина конуса отводится в сторону и просвет трахеи открывается. Закрывание и открывание дыхалец обеспечивают более интенси вный обмен газов при дыхательных движениях насекомого. При закрытом дыхальце полностью прекращается доступ воздуха в трахею, что позволяет пчелам длительное время находиться в отравленном воздухе, изолировав себя от него.
Регуляция работы дыхалец основана главным образом на рефлекторном торможении и возбуждении мышц, обслуживающих замыкающий аппарат. Помимо этого центрального механизма регуляции действует и периферический механизм, обусловленный блокирующим влиянием диоксида углерода на нервно-мышечную передачу. Дыхательный замыкающий аппарат реагирует на изменения концентрации диоксида углерода. При высокой концентрации углекислоты нервно-мышечная передача полностью блокируется и дыхальца вообще не закрываются.
Отходящие от дыхалец короткие трахейные стволы ведут в воздушные мешки. Одна пара самых крупных воздушных мешков расположена симметрично по бокам брюшка. В груди расположены четыре парных и два непарных мешка. Вмешках отсутствуют спиральные утолщения, поэтому их стенки могут опадать при сжатии брюшка и увеличиваться при его расширении. В голове и груди воздушные мешки не могут спадаться, как в брюшке, так как они постоянно заполнены воздухом и сохраняют свой объем. Стигмы дыхательной пластинки жалоносного аппарата также соединены с небольшими воздушными мешками, которые дают отростки в большие воздушные мешки. Воздушные мешки брюшка, груди и головы соединены друг с другом трахейными стволами.
Воздушные мешки обеспечивают интенсивное дыхание пчелы во время полета, когда необходим добавочный кислород в связи с повышенным уровнем обмена веществ. Кроме того, воздушные мешки участвуют в вентиляции крыловых мышц и регулируют объем внутренних органов (кишечника, яичника), что в значительной степени облегчает полет пчелы. От воздушных мешков отходят многочисленные трахеи. Диаметр трахей постепенно уменьшается.
Мельчайшие трахеи с просветом 1мкм и менее называют трахеолами. Они лежат непосредственно на поверхности клеток, но иногда проникают внутрь их и близко подходят к митохондриям. Некоторые трахеи, имеющие диаметр до 2-5 мкм, заканчиваются ос обой трахейной (звездчатой) клеткой. Стенки трахеол и трахейных клеток проницаемы для газов и воды. Концы их всегда замкнуты, поэтому обмен газов происходит через их стенки. Эти клетки принимают участие в контроле снабжения тканей кислородом.
ПРОЦЕСС ДЫХАНИЯ
При дыхании воздух через дыхальца проникает в главные трахейные стволы и далее по разветвлениям трахей меньшего диаметра достигает трахеол, через которые кислород поступает к тканям и клеткам.
Большинство насекомых совершают дыхательные движения, заключающиеся в ритмичных сжатиях и расширениях брюшка. Так, брюшко пчелы во время таких движений увеличивается в длину на 12%. в ширину на 2,5%. Движения брюшка обеспечивают механическую вентиляцию трахейной системы. При расширении брюшка давлениевнутри воздушных мешков значительно снижается, что ведет к засасыванию воздуха через дыхальца. Объем воздуха в воздушных мешках и трахейных стволах при интенсивном дыхании увеличивается и уменьшается на 1/3 их общего объема. Ритм дыхательных движений, а следовательно и интенсивность трахейной вентиляции, зависит от внешних условий и состояния пчелы. Так, спокойно сидящая пчела совершает 40 дыхательных движений в минуту, вернувшаяся из полета — до 150. Наиболее интенсивны дыхательные движения у пчелы во время полета.Установлено, что вентиляция трахейной системы у пчелы носит направленный характер — воздух входит в одни дыхальца и выходит в другие. Воздух в тело пчелы попадает через брюшные дыхальца в момент расслабления брюшка. При сжатии брюшка воздух перегоняется из брюшка в грудь. Брюшные дыхальца в этот момент закрыты, а проподемальные открыты. Отработанный воздух выходит через проподемальные дыхальца. При полете воздух поступает в трахейные стволы через брюшные и первое грудное дыхальца, а выходит через проподемальные дыхальца.
Трахейная система вентилируется еще дополнительно за счет мышц при полете. Воздушные мешки в голове вентилируются за счет изменений кровяного давления: при повышении его воздушные мешки сжимаются, выталкивая воздух, при снижении — раздуваются, засасывая воздух.
При выдохе воздушные мешки брюшка опадают, что создает хорошие условия для отсасывания воздуха из трахей в мешки во время выдоха. Тем самым воздушные мешки выполняют роль резервуаров, в которых воздух при вдохе накапливается, а затем при выдохе, главным образом во время фазы сжатия, несколько продвигается в трахеи.
С фазой сжатия связана ритмическая работа дыхалец, которые открываются и закрываются синхронно с общими дыхательными движениями пчелы. Ритмическая работа дыхалец и дыхательные движения брюшка регулируются как нервным, так и химическим путем.
В регуляции дыхательных движений принимают участие два типа нервных центров. Они определяют ритм и амплитуду сокращений туловищных мышц. Первичный дыхательный центр вызывает дыхательные движения, а вторичный стимулирует или тормозит деятельность первичного центра.
Первичные центры соответствуют отдельным ганглиям брюшка и заднегруди. Кажды й из них контроли рует дыхательные движения своего сегмента, но один из ганглиев является ведущим, который определяет ритм дыхательных движений всего тела. В качестве ведущего дыхательного центра выступает первый брюшной ганглий. Вторичные дыхательные центры соответствуют головным и грудным ганглиям. Импульсы этих ганглиев могут изменить ритм дыхательных движений, зарождающихся в первичных центрах. Вторичные дыхательные центры способны реагировать на изменение рН гемолимфы, которое определяется уровнем содержания диоксида углерода и недоокисленных продуктов обмена веществ.
Повышенное содержание СО2 приводит к увеличению частоты нервных импульсов во вторичном центре и открыванию дыхалец на длительное время. Недостаток кислорода также стимулирует дыхательные движения брюшка пчелы.
С помощью дыхательных движений при открытых дыхальцах воздух легко проникает в крупные трахеи. Проникновение же его в тонкие трахеи и трахеолы путем нагнетания невозможно вследствие огромного капиллярного сопротивления. В этом случае, согласно диффузионной теории А. Крога (1920), транспортирование О 2 и СО 2 по трахейной системе осуществляется благодаря процессам газовой диффузии. Движущая сила диффузии — разность парциальных давлений газа в атмосфере и в концевых разветвленных трахеях. Парциальное давление кислорода выше в атмосфере и ниже в тканях, испытывающих потребность в этом газе. В результате возникают диффузионные потоки кислорода, направленные внутрь тела насекомого.
Вентиляция трахейной системы обеспечивает не только поступление в организм кислорода, но и удаление из него диоксида углерода. Это достигается как при дыхательных движениях (путем выдыхания), так и с помощью диффузии через кожу. Последний способ имеет немаловажное значение ввиду того, что диффузия диоксида углерода через ткани совершается в 36 раз интенсивнее, чем кислорода. Вместе с тем б олыпая часть диоксида углерода удаляется через трахейную систему в газообразном состоянии и до 25% его выводится из организма диффузно.
При сравнении строения дыхательной и кровеносной систем отмечается взаимная отрицательная связь между ними: кровеносная система медоносной пчелы развита слабо и, за исключением аорты, никаких кровеносных сосудов не имеет; напротив, трахейная дыхательная система сильно развита, мельчайшие разветвления ее проникают в самые отдаленные части тела. Такое различие в строении дыхательной и кровеносной систем определяется их функциями: кровеносная система в основном разносит по телу питательные вещества и выво дит продукты распада, а дыхательная система выполняет функцию газообмена, доводит до клеток тела кислород, обеспечивая тем самым процесс окисления, и удаляет из тела продукт окисления — СО2.
Так как при дыхании поглощаемые и выделяемые вещее тва находятся в газообразном состоянии, процесс дыхания называют также газообменом. В обменных процессах, происходящих в организме, большое значение имеет дыхательный коэффициент — отношение количества выделенного СО2 к количеству поглощенного кислорода. Вычисление дыхательного коэффициента позволяет установить, какие вещества используются для окисления во время дыхания пчелы. При полном окислении углеводов он равенЬ белков — 0,78-0,82, жиров — 0,7.
Значение дыхательного коэффициента зависит от возраста пчел и используемого корма. Так, у молодых пчел, выполняющих функции по кормлению личинок и усиленно потребляющих белковый корм (пыльцу), дыхательный коэффициент равен 0,7; улётных пчел, питающихся главным образом сахаром, в состоянии покоя и при непродолжительном полете — 1; у пчел, использующих для питания преимущественно мед (взимний период), — 0,94. Кроме того, на этот показатель влияет температура наружного воздуха.
Максимального значения он достигает при 17 °С, а минимального —при 32 °С.
К внешним факторам, влияющим на газообмен и потребление кислорода, относится влажность. Вусловиях пониженной влажности, когда возникает угроза быстрого испарения воды из организма, насекомые часто закрывают дыхальца. Это приводит к снижению интенсивности дыхания.
Потребление пчелами кислорода резко повышается с изменением их активности. Так, у летящей пчелы по сравнению с находящейся в полном покое поглощение кислорода увеличивается в 48раз и более.
Потребность пчелиной семьи в кислороде складывается из потребности самих пчел и их ра сплода, находящегося в разных стадиях развития. Установлено, что в нормальном состоянии для семьи пчел (вместе с расплодом) требуется в активный летний период около 20л воздуха в час (на 1 кг пчел), а зимой — лишь около 4 л.
Особенно велика потребность пчелиной семьи в кислороде во время выполнения интенсивных работ по выращиванию расплода, выделению воска, строительству сотов, переработке нектара в мед.
На интенсивность обмена веществ и потребление кислорода огромное влияние оказывает состояние пчелиной семьи. Беспокойство пчел, вызванное любым фактором, резко повышает обмен веществ.