

О.А. ВЕРХОВСКИЙ, Ю.Н. ФЕДОРОВ
ВИЭВ
М.М. ГАРАЕВА, Т.И. АЛИПЕР
НПО НАРВАК
Основная задача промышленного птицеводства - обеспечение сохранности и продуктивности птицы. Она напрямую связана с технологией кормления и содержания, программой вакцинации, применением химиотерапевтических препаратов, генотипом птицы, мерами санитарно-биологического контроля. При этом важнейшим фактором, влияющим на эпизоотическое благополучие птицеводческих хозяйств, высокую продуктивность и устойчивость особей к неблагоприятным факторам окружающей среды, является состояние иммунной системы.
Иммунная система птиц филогенетически более ранняя, чем таковая млекопитающих, и ее структурные элементы (первичные и вторичные лимфоидные органы и ткани, лимфоциты, макрофаги, цитокины, система комплемента и др.), так же как и механизмы формирования иммунитета, имеют некоторые отличительные особенности. В частности, это касается структурных и функциональных свойств иммуноглобулинов (Ig) - белков организма, секретируемых В-лимфоцитами и являющихся основным компонентом системы гуморального иммунитета.
У зародыша птиц предшественники В-лимфоцитов образуются в желточном мешке, а впоследствии - в костном мозге, затем они мигрируют в фабрициеву сумку, где происходят их окончательное созревание и дифференцировка. Далее В-лимфоциты попадают во вторичные лимфоидные органы и ткани (селезенка, костный мозг, Гарднерова железа, лимфоидные узелки, лимфоидная ткань, ассоциированная со слизистыми оболочками бронхов, кишечника и конъюнктивы) - основное место формирования специфического иммунного ответа.
У птиц, как и у млекопитающих, антигенраспознающие рецепторы Т- и В- лимфоцитов происходят от общего филогенетического предшественника и принадлежат к суперсемейству иммуноглобулинов. Все рецепторы на одной клетке идентичны, поэтому она способна связывать и отвечать только на один эпитоп чужеродного антигена. В процессе иммунного ответа В-клетки секретируют в кровоток растворимые молекулы lg-рецептора, которые обладают определенной специфической активностью и называются антителами. Таким образом, иммуноглобулины функционируют в качестве антигенраспознающих рецепторов, а также в виде циркулирующих антител, присутствующих в биологических жидкостях организма.
Основную структурную единицу молекулы Ig образуют 2 одинаковые тяжелые (Н) и 2 одинаковые легкие (L) полипептидные цепи, которые при помощи дисульфидных мостиков и нековалентных связей формируют Y-образную молекулу, существующую в трехмерной форме. Тип тяжелой цепи определяет класс (или изотип) иммуноглобулина, а легкие, общие для всех изотипов, участвуют в формировании и поддержании структуры антигенсвязывающих участков молекул Ig. У птиц в отличие от млекопитающих существует только один тип L-цепей -Х [18].
В мономерной форме молекула иммуноглобулина (H2L2) двухвалентна, так как содержит 2 идентичных антиген-связывающих участка, которые могут взаимодействовать с 2 эпитопами одинаковых или разных молекул антигена. В состав антигенсвязывающих участков, детерминирующих специфичность антительного ответа, входят 2 вариабельных (V) домена, однако эффекторная активность антител обусловлена изотипом их тяжелой цепи, входящей в состав константного (С) домена.
У птиц в отличие от млекопитающих обнаружено и описано всего 3 изотипа иммуноглобулинов: IgY (IgG), IgM и IgA (см. таблицу). IgY (IgG) доминирует в сыворотке крови и яичном желтке, IgA - в секретах, яичном белке и желчи, a IgM - в сыворотке крови и яичном белке [6, 8, 13, 28].
Наиболее изучены Ig кур (отряд курообразные - Galliformes) и уток (отряд гусеобразные - Anseriformes).
Иммуноглобулины класса Y (G). Первоначально основной класс иммуноглобулинов в сыворотке крови птиц назвали IgG, однако обнаруженные G.A. Leslie и LW. Clem (1969) значительные физико-химические и структурные различия между IgG млекопитающих и птиц, позволили переименовать его в иммуноглобулины IgY [17]. Обоснованность этого термина подтвердили результаты молекулярного клонирования генов, кодирующих каждый изотип Ig, показавшие, что тяжелые (v) цепи IgY птиц содержат 5 доменов: один вариабельный (VH) и четыре константных (СН1 - СН4), в то время как
Основные физико-химические и биологические свойства иммуноглобулинов кур

тяжелые (у) цепи IgG млекопитающих состоят только из 4 (VH и СН1 - СН3) [10, 24, 29]. В молекуле IgY нет шарнирной области, которая есть между СН1 и СН2 доменами в молекуле IgG, что делает v-цепи менее мобильными по сравнению с у-цепями, и в конечном итоге оказывает влияние на их биологическую активность [28]. Таким образом, больший размер Н-цепи IgY птиц обусловливает ее большую по сравнению IgG млекопитающих молекулярную массу (68 кД и 50 кД соответственно). Физико-химические свойства IgY варьируют у разных видов птиц. Так, IgY кур способен преципитировать антиген только в солевом растворе высокой концентрации (1,5 М/л) или при кислом рН (< 5,0), а период полураспада составляет 4,1 дня [11, 14]. Предполагают, что молекула IgY кур более гидрофобна, чем IgG млекопитающих [7].
У кур IgY функционирует в мономерной форме с коэффициентом седиментации 7.8S, хотя I. Tizard Г°Я] общает о наличии полимерной фс - '(19S), присутствующей в сыворотк рови старых особей. Кроме того, у некоторых видов птиц (в частности, у уток и гусей) помимо полноразмерного IgY обнаружили его усеченную изоформу с коэффициентом седиментации 5,7S, названную IgY (AFc), H-цепь которой содержит только 3 домена (VH, CH1 и СН2) [18, 19]. Когда молекула IgY (AFc) продуцируется В-клетками, то происходит блокировка транскрипции СН3 и СН4 эксонов, что приводит к отсутствию Fc-области, определяющей биологические свойства молекулы Ig [28]. Таким образом, эффекторные функции IgY (AFc) остаются непонятными.
Результаты последних исследований показали, что IgY птиц - общий предшественник IgG и IgE млекопитающих, а его основные биологические функции сочетают таковые этих 2 изотиповIg[8, 18, 26, 29]. Антитела lgY-изотипа у птиц, так же как и IgG млекопитающих, являются доминирующими системными антителами, вовлеченными в процессы нейтрализации вирусов, опсонизации бактериальных патогенов, активации системы комплемента и материнского (трансовариального) иммунитета. Поэтому, исходя из общих биологических свойств Ig этих изотипов, многие авторы при описании иммуноглобулинов птиц используют название IgG вместо IgY [3, 21, 26]. С другой стороны, структура IgY больше похожа на таковую IgE, чем на IgG, а антитела данного изотипа, присутствуя в тканях, участвуют в реакции, подобной анафилактической у млекопитающих [9].
Помимо птиц антитела lgY-изотипа обнаружены у некоторых видов рыб, амфибий и рептилий [28].
Иммуноглобулины класса М. У всех позвоночных филогенетически самые ранние и первые антитела, появляющиеся в процессе иммунного ответа, относятся к lgM-изотипу. Структура и биологические свойства IgM кур подобны таковым млекопитающих. Они существует в 2 формах: в виде полимера, представляющего большую молекулу, состоящую из пяти Ig-cyбъединиц (М.м. 180 кД каждая), соединенных J-цепью, и в мономерной форме, которая связана с клеточной поверхностью В-лимфоцита и выступает в роли антигенного рецептора. Полимерный IgM количественно охарактеризован в сыворотке крови кур, а также он обнаружен в желчи и слизистой кишечника. Мономерный IgM в очень небольшом количестве присутствует в желтке и сыворотке крови вылупившихся цыплят [6]. Поскольку молекула IgM теоретически может иметь 10 антигенсвязывающих участков, то антитела данного изотипа более эффективны в реакции агглютинации [28].
Иммуноглобулины класса А. У цыплят IgA существует в мономерной и димерной формах, а в секретах - в полимерной форме, связанной с секреторным компонентом (slgA). Структура мономера IgA у цыплят подобна таковой у млекопитающих, однако его тяжелые цепи содержат четыре Сн домена, в то время как а-цепи млекопитающих состоят только из трех с шарнирной областью вместо Са2 домена [19, 20].
У птиц, как и у млекопитающих, мономер IgA присутствует в сыворотке крови, в то время как slgA в количественном отношении преобладает в секретах респираторного и кишечного трактов, где составляет основу местного иммунитета (иммунитета слизистых) против бактерий и вирусов. Кроме slgA во всех секретах кур обнаружен свободный секреторный компонент, а в секрете респираторного тракта - большое количество IgY [16].
Желчный пузырь считают важным лимфоидным органом, в котором через 3-8 дней после вылупливания цыплят формируется лимфоидная ткань - место синтеза и секреции не только slgA, но и IgY, и IgM [28, 30]. При проникновении антигена в желчный пузырь начинается синтез специфических антител, которые затем выявляют в желчи и сыворотке крови. IgA иногда присутствует в желчи, кроме сыворотки крови и секретов, его выявляют в яичном белке.
Иммуноглобулины желтка. По разным данным, желток куриного яйца содержит IgY в высокой концентрации (8 -20 мг/мл) [4, 22]. Это обусловлено легким переходом сывороточных антител в белок яйца находящегося в яичнике, далее происходит активный перенос и аккумуляция IgY в желточном мешке. В практическом аспекте такие свойства IgY, как его высокая концентрация в желтке, ревматоидный фактор, неспособность связывать белки А и G, активировать систему комплемента и интерферировать с IgG млекопитающих, привели к разработке так называемой lgY-технологии - альтернативе традиционному методу получения поликлональных антител на животных [23, 27]. В настоящее время lgY-антитела различной специфичности, полученные гипериммунизацией кур и выделенные из желтка, используют в диагностических и лечебных целях в ветеринарии и медицине [5, 25]. Установлено, что питьевой йогурт, содержащий 1% lgY-антител куреазе Helicobacter pylori, можно эффективно применять для лечения больных детей вместо антибиотикотерапии [15].
Материнские IgY из желточного мешка начинают передаваться зародышу в первую неделю эмбриогенеза, а пик его приходится на последние три дня перед выведением цыпленка. Помимо IgY зародыш получает в небольшом количестве материнские IgM и IgA, диффундирующие в амниотическую жидкость. Выведенные цыплята способны абсорбировать Ig желтка еще в течение 3 дней, и в это время содержание их сывороточных IgY (5-10 мг/мл) превышает показатель взрослой птицы. Пассивно приобретенные антитела подвергаются катаболизму, и через 2 -3 нед их уровень составляет менее 1 мг/мл [28]. Пассивная передача антител от курицы через яйцо цыплятам - важнейший фактор иммунитета против инфекционных болезней в первые дни их жизни. Оценка уровня антител. В настоящее время существует стройная методология оценки клеточных факторов иммунитета млекопитающих, и прежде всего человека. Однако для птиц подобные тест-системы отсутствуют, поэтому определение уровня специфических антител остается в большинстве случаев единственным средством оценки иммунологического состояния организма. Выявляемые в иммунохимических тестах антитела, по своей природе можно подразделить на материнские, постинфекционные и поствакцинальные, поэтому практическое значение имеют: уровень трансовариальных антител у цыплят в первые дни (недели) жизни; напряженность иммунитета и иммунологическая эффективность вакцинных препаратов; ретроспективная диагностика и эпизоотологический мониторинг инфекции.
Определение уровня трансовариальных антител у цыплят в первые дни (недели) жизни имеет большое значение. Известно, что активная вакцинация, проводимая на фоне высокого содержания материнских антител, не обеспечивает формирования напряженного иммунитета вследствие нейтрализации колостральными антителами вакцинного вируса. В то же время если с вакцинацией запоздали, то на фоне исчезновения материнского иммунитета цыпленок может быть инфицирован вирулентным возбудителем или у него может возникнуть гиперчувствительная поствакцинальная реакция. Таким образом, существует определенный промежуток времени ("окно восприимчивости") между возрастом, когда цыпленок становится чувствительным к заражению вирулентным вирусом и таковым, когда он должен быть вакцинирован. Для того чтобы свести этот временной показатель до минимума, специалистам хозяйств необходимо разработать стратегию вакцинации поголовья с учетом корректировки сроков ее начала и последующего периода формирования иммунитета. Только в этом случае эффект от применения вакцин будет максимальным.
Оценивая напряженность иммунитета и иммунологическую эффективность вакцинных препаратов, изучают динамику появления, нарастания и длительность сохранения поствакцинальных антител, что опосредованно является маркером напряженности иммунитета. Сейчас это особенно актуально, так как практическая ветеринарная служба использует огромное количество разных типов отечественных и зарубежных вакцин, в том числе и поливалентных, которые должны обеспечить формирование напряженного иммунитета одновременно против нескольких патогенов. Однако при их разработке и применении на восприимчивом поголовье необходимо учитывать возможность интерференции антигенов, входящих в состав вакцины, в организме птицы в процессе иммуногенеза. Зачастую одна серия исследований по оценке гуморального иммунитета в динамике или на разновозрастном поголовье может помочь решить сразу обе задачи (см. рисунок).
Для ретроспективной диагностики и эпизоотологического мониторинга
инфекции необходимо регулярно определять уровень специфических антител в сыворотке крови неиммунного поголовья, что позволяет судить о циркуляции конкретного возбудителя в хозяйстве или на обследуемой территории, а также своевременно принимать адекватные меры при ухудшении эпизоотической ситуации.
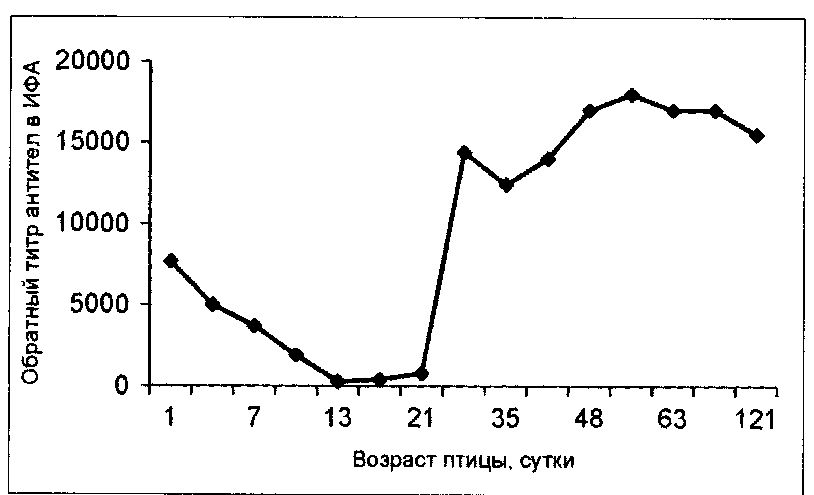
Динамика содержания IgY (1дв)-антител к вирусу инфекционной бурсальной болезни в сыворотке крови кур разных половозрастных групп, вакцинированных в плановом порядке в 9-, 19- и 100-дневном возрасте. Приведены средние значения показателей. Для исследования использовали соответствующие ИФА-наборы производства НППАВИВАК[2].
В последние годы для проведения подобных исследований помимо традиционных серологических тестов (реакция нейтрализации вируса, реакция торможения гемагглютинации) широко используют иммуноферментный метод. Его высокая чувствительность и специфичность, доступность и стабильность реагентов, экспрессность, возможность инструментального учета результатов и автоматизации процесса при массовых исследованиях привели к появлению на рынке коммерчески доступных отечественных и зарубежных ИФА-наборов, применяемых главным образом для выявления специфических lgY-антител в сыворотке крови и яичном желтке [1].
Динамика формирования гуморального иммунного ответа у птиц подобна таковой у млекопитающих [3, 10, 28, 29].
Заключение. Всестороннее изучение иммуноглобулинов птиц, их количественный анализ и оценка распределения изотип-специфических антител дают важную информацию об иммунологическом состоянии организма. Решение проблемы, связанной с изучением природы и биологических функций Ig различных изотипов птиц, представляет интерес не только для фундаментальной ветеринарной иммунологии, но и способствует дальнейшему изучению инфекционных болезней, разработке и совершенствованию методов иммунодиагностики, средств терапии и специфической профилактики.
ЛИТЕРАТУРА
1. Верховский О.А. и др. // Био. 2004. № 5 (44).
2. Верховский О.А. // Мат. Юбилейной научно-практической конференции "Белые ночи 2005".- СПб. 2005.
3. Al-Garib S.O. etal. //Avian Dis. 2003. 47(1).
4. CamenischG. et al.//FASEB J. 1999. 13.
5. Carlander D. et al. // Immunol. Res. 2000. 21.
6. Dahan A. et al. // Nucleic Acids Res. 1983. 11.
7. Davalos-Pantoja L. et al. // Biomater. Sci. Polym. Ed. 2000. 11.
8. Diseases of poultry. 11th Edition. / Ed. Y.M.Saif, Iowa State Press, 2003.
9. Faith R.E. et al.//Immunology. 1973.25.
10. Foppoli J.M. etal.//J. Immunol. 1978. 120.
11. Gallagher J.S. et al. // Immunochemistry. 1970. 7.
12. Garcia D.A. et al. //Biomedica. 2005. 25(4).
13. Goudswaard J. etal. // PoultrySci. 1977.56.
14. HigginsD.A.//Res. Vet. Sci. 1976. 21.
15. Horie K. et al. // J. Dairy Sci. 2004. 87.
16. Karaca K. et al. // Hybridoma. 1997. 16.
17. Leslie G.A. et al. // J. Exp. Med. 1969. 132.
18. Lundqvist M.L etal.//Dev. Сотр. Immunol. 2006.30(1-2).
19. MagorK.E. etal. //J. Immunol. 1994. 153.
20. MansikkaA.//J. Immunol. 1992. 149.
21. Ratcliffe M.J.H.// Dev. Сотр. Immunol. 2006. 30 (1-2).
22. Ruan G.R et al. // Protein Expr Purif. 2005. 44 (1).
23. Schade R. et al.//Altern. Lab. Anim. 2005.33(2).
24. Sharma J.M. //ActaVet. Hung. 1997. 45.
25. Shin J.H. etal.//Clin. Diagn. Lab. Immunol. 2002. 9 (5).
26. Suzuki N. etal. // Glycobiology. 2004. 14 (3).
27. Tini M. et al. // Сотр. Biochem. Physiol. - Part A: Molecular & Integrative Physiology. 2002. 131(3).
28. Tizard I. // Seminars in Avian and Exotic Pet Medicine. 2002. 11.
29. WarrG.W. etal.//Immunol. Today. 1995. 16.
30. WatanabeH. et al.//J. Immunol. 1975. 115.
журнал "Ветеринария" №11 2007