
Монгалёв Н.П., Борисенков М.Ф.
Федеральное государственное бюджетное учреждение науки Институт физиологии Уральского отделения РАН,
В формировании структур и функционировании репродуктивных органов у раз-личных видов значительная роль принадлежит иммунной системе. Иммуноциты обеспечивают стерильность полового трак-та, селекцию половых клеток, а также процесс оплодотворения и развития плода [5]. Имея рецепторы к половым гормонам, они участвуют в процессе овуляции доминантного фолликула и функционировании желтого тела. Наличие в эозинофилах рецепторов эстрогенов объясняет присутствие этих клеток при формировании желтого тела и их инфильтрацию в ткани матки в предовуляторный период [10]. Определена роль нейтрофильных лейкоцитов в регуляции целостности плодных оболочек [6] и участие моноцитов на ранних стадиях функциональной регрессии желтого тела в яичнике у коров [9].
Показано, что в крови коров из сосудов яичников и матки по сравнению с яремной веной уровень половых гормонов на 2-3 порядка выше, наряду с многократно увеличенным количеством лимфоцитов, что дало основание обозначить это явление как физиологический лимфоцитоз [1]. В то же время отсутствуют работы по исследованию реактивности иммуноцитов в крови из сосудов репродуктивных органов в течение полового цикла у животных в пубертатный и лактационный периоды.
Целью данной работы явилось определение физиологической значимости лейкоцитоза в распределении функционально разграниченных лейкоцитов у телок и коров вблизи источников половых гормонов - в яичниковых и маточных сосудах репродуктивных органов в лютеиновую и фолликулиновую фазы полового цикла.
Материал и методы исследования
Исследования проведены на клинически здоровых телках и коровах холмогорской породы 1-6 отелов (п=21) со средней продуктивностью 4000 кг за лактацию в зимне-весенний период на ферме Коми научного центра УрО РАН. Рацион был сбалансирован по основным питательным веществам с учетом физиологического состояния и уровня молочной продуктивности животных согласно нормам ВИЖа. Длительность полового цикла, протяженностью в среднем 19-21 день, фиксировали как период между двумя овуляциями. Для исследования использовали гепаринизированную кровь, взятую из яремной вены у тёлок и коров 1-6-го отелов, а также из яичниковых и маточных вен у животных того же возрас-та сразу после их убоя на специальном пункте (п=24) [2]. Лейкоформулу диф-ференцировали при анализе 300 клеток. Полученные результаты обрабатывали статистически с использованием пакета прикладных программ Windows (Basic). Достоверность различий оценивали по t-критерию Стьюдента при уровнях значимости от 0,05 до 0,001.
Результаты и их обсуждение
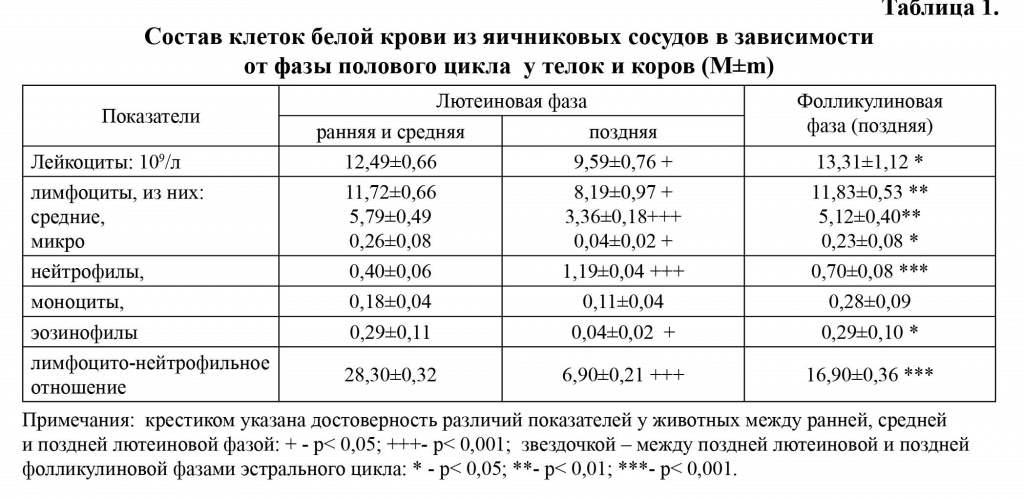
Лейкоциты. В крови из сосудов репродуктивных органов по сравнению с яремной веной у телок и коров отмечали повышение уровня лейкоцитов, уменьшение количества лейкоцитов в крови из сосудов репродуктивных органов в позднюю лютеиновую фазу полового цикла (р<0,05) и их увеличение в позднюю фолликулиновую фазу (р<0,05) (табл. 1).
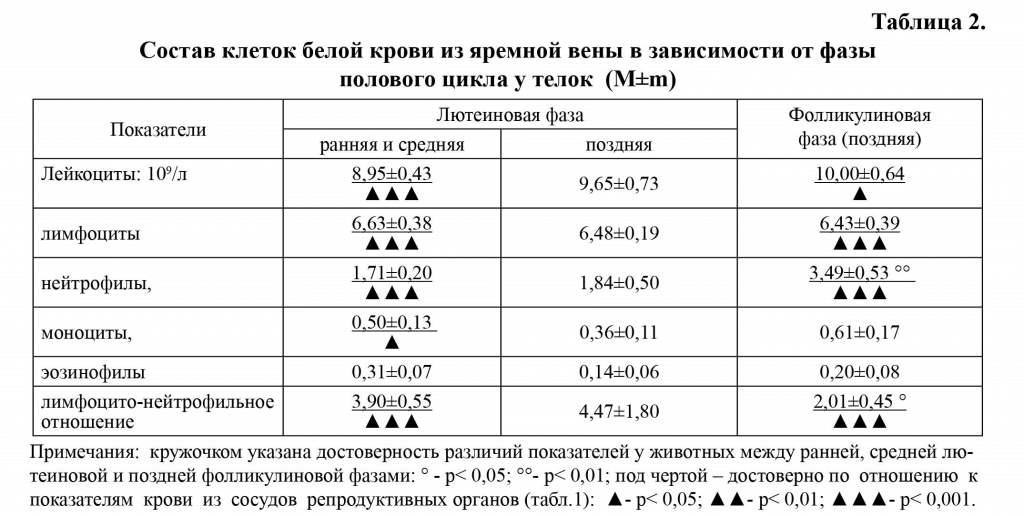
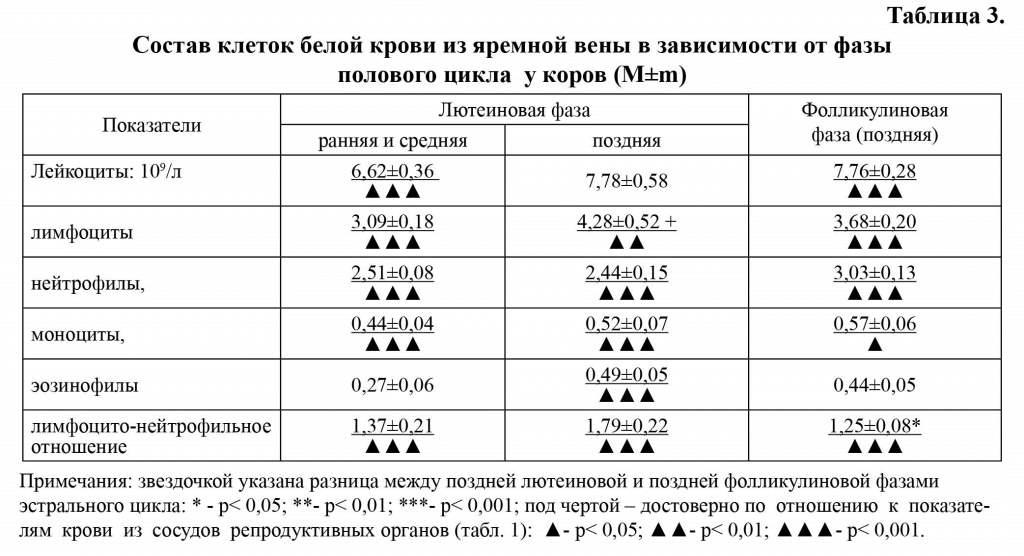
В крови из яремной вены у телок и коров определилась тенденция к увеличению количества лейкоцитов в позднюю фолли-кулиновую фазу по отношению к ранней и средней лютеиновой фазами цикла (табл. 2,3).
Таким образом, несмотря на уменьшение содержания лейкоцитов в крови у коров с возрастом, их количество оставалось на высоком уровне в крови яичниково-ма-точных сосудов в течение всех исследуе-мых периодов полового цикла.
Лимфоциты. Динамика лимфоцитов в разные периоды полового цикла соответствовала таковой общему количеству лейкоцитов (табл. 1,2,3). Заметно уменьшилось количество лимфоцитов в крови яичниковых сосудов у животных в позднюю лютеиновую фазу по сравнению с ранней и средней лютеиновой фазами цикла (р<0,05). Это уменьшение сказалось на количестве разных по морфофункци-ональным свойствам субпопуляций лим-
фоидных клеток: средних лимфоцитов (р<0,001) и микролимфоцитов (р<0,05) способных в норме к бласттрансформа-ции. В позднюю фолликулиновую фазу полового цикла по сравнению с поздней лютеиновой фазой количество лимфоцитов у коров вновь увеличилось (р<0,01). С возрастом животных распределение количества лимфоидных клеток по фазам цикла изменялось таким образом, что у телок в позднюю лютеиновую фазу этот показатель сохранялся на одном уровне, тогда как у коров в этот период полового цикла представлял наибольшую величину (р<0,05).
Нейтрофилы. Количество нейтрофилов в крови из яичниковых сосудов меньше, по сравнению с содержанием их в яремной вене телок и коров в разные периоды эстрального цикла (табл. 1,2,3). В крови сосудов репродуктивных органов отмечали их абсолютное увеличение в позднюю лютеиновую фазу (р<0,001) и, напротив, у телок в магистральных сосудах количество нейтрофилов преобладало в позднюю фолликулиновую фазу цикла (р<0,001). Умень-
шение количества нейтрофилов в яичниково-маточном кровотоке можно объяснить, по-видимому, тем, что эти клетки, особенно крупные, отличаются чрезвычайно высокой функциональной мобильностью, поскольку они имеют экспрессированные рецепторы к хемокинам, которые секретируются эндотелием сосудов и активно мигрируют из кровеносного русла в ткани, обладая наиболее высокой чувствительностью к хемотаксису [3].
Лимфоцито-нейтрофильное отношение. Изменения количества лимфоцитов и нейтрофилов в разные фазы полового цикла в крови сосудов репродуктивных органов и яремной вены у животных не совпадают, что обусловило вариабельность Л/Н отношения (табл. 1-3). Минимальное Л/Н отношение в крови яичниково-маточных вен отмечали в позднюю лютеиновую фазу полового цикла по сравнению с ранней и средней лютеиновой фазой (р<0,001) и его повышение в позднюю фолликулиновую фазу цикла (р<0,001). В крови из яремной вены у телок и коров в позднюю лютеиновую фазу по сравнению с ранней и средней лютеиновой фазой проявилась тенденция к увеличению Л/Н отношения с последующим уменьшением этого показателя в позднюю фолликулиновую фазу цикла (р<0,05).
Уменьшение Л/Н отношения у телок за 1-2 дня до овуляции и увеличение Л/Н от-ношения у коров за 4-5 дней до овуляции явились маркерными показателями адекватного взаимодействия иммунной и ре-продуктивной систем при формировании полноценного эстрального цикла. Время, в течение которого осуществлялся переход от максимального Л/Н отношения к минимальному Л/Н отношению в эстральном цикле животных, может соответствовать длительности фолликулиновой фазы, которая у коров продолжительнее, чем у телок.
Эозинофилы и моноциты. Наибольшее количество эозинофилов и моноцитов в крови яичниковой и яремной вен у телок и коров определяли в раннюю и среднюю лютеиновую фазу полового цикла, а наименьшее - в позднюю лютеиновую (табл. 1-3). В крови из яремной вены телок содержание моноцитов выше, чем в крови со-судов репродуктивных органов в раннюю и среднюю лютеиновую фазу (р<0,05), тогда как количество эозинофилов находилось на одном уровне. Количество моноцитов и эозинофилов у животных в сосудах крови репродуктивных органов в позднюю лютеиновую фазу цикла уменьшилось на 39%, тогда как в крови яремной вены на 28 и 14,3% соответственно.
Мобильность эозинофилов и моноцитов, как и нейтрофилов, которые обладают высокой хемотактической чувствительностью [3], находилась в зависимости от уровня половых гормонов. В крови оттекающей от яичника с доминантным фолликулом, где преобладает продукция эстрадиола над прогестероном, наблюдали более высокое содержание моноцитов и эозинофилов [1].
Таким образом, на основании представления об избирательной инфильтрации лейкоцитов в окружающие ткани можно интерпретировать повышение активности лимфоцитов, моноцитов и эозинофилов в позднюю лютеиновую фазу цикла, а нейтрофилов - в крови яичниково-маточных сосудов в фолликулиновую и раннюю и среднюю лютеиновую фазы цикла у телок и коров. Эти изменения соответствовали массовой атрезии фолликулов в позднюю лютеиновую фазу. Поэтому переход эстрогенов из атретических фолликулов в кровь, вероятно, стимулировал пролиферацию клеток белой крови.
В то же время клеточный состав белой крови в яичниково-маточных сосудах из-менялся не всегда (строго) в соответствии с гормональной активностью яичников, поскольку, возможно, эндотелий и ткань матки, активированная гормонами, способна выделять факторы (хемоатрактанты) не менее активно влияющие на миграцию лейкоцитов, чем половые стероиды [1]. Определенное значение в распределении лейкоцитарного пула, по-видимому, имел локальный яичниково-маточный кровоток. В среднюю и позднюю лютеиновую фазу цикла кровь текла в сторону яичников, а в фолликулиновую и раннюю лютеиновую фазы - в сторону матки [8].
Поддержание состояния физиологического лейкоцитоза в крови яичниково-маточных сосудов у животных, вероятно, способствовало ускоренному использованию функционально разграниченных клеток белой крови для формирования структуры репродуктивных органов, при этом изменение Л/Н отношения в зависимости от фазы эстрального цикла являлось одним из индикаторов оптимального взаимодействия иммунной и репродуктивной систем.
С началом беременности физиологический лейкоцитоз в крови сосудов репродуктивных органов коров нивелировался [1], что, вероятно, обусловлено избирательной активацией гормональных регуляторов, прямое воздействие которых на органы и ткани иммунной системы обеспечивалось рецепторно-опосредованным путем [7]. В первые дни беременности снижалась реактивность фагоцитарного звена иммунитета также со стороны материнской иммунной системы [6].
При гипофункции яичников физиологический лейкоцитоз отсутствовал, в сосудах репродуктивных органов лейкоцитарный пул в крови по клеточному составу при-ближался к таковому из яремной вены [1]. Можно предположить, что гипофункция яичников у коров характеризовалась снижением гормонпоэтической функции, то есть отсутствием циклического подъема синтеза гормонов. Нарушение стероидогенеза происходило, по-видимому, на самом раннем его этапе - в период превращения холестерина в прогненолон, который контролировался лютеинизирующим гормоном гипофиза [4]. Низкая продукция прогестерона и эстрадиола не обеспечивали поддержание высокого уровня лимфоидных клеток в крови сосудов яичников и матки, в том числе и достаточной бласттрансформации, которая характерна для крови коров.
Заключение
Периодичность перераспределения количества функционально разграниченных лейкоцитов в яичниковых и маточных сосудах репродуктивных органов телок и ко-ров в течение полового цикла реализуется на уровне физиологического лейкоцитоза (лимфоцитоза) обусловленного высоким уровнем содержания в крови половых гормонов.
Сравнительный анализ содержания различных типов лейкоцитов крови из яремной и яичниковой вен у телок и коров показал проявление асинхронности изменения клеточного состава белой крови в зависимости от фазы полового цикла у животных. На основании представления об избирательной инфильтрации лейкоцитов из крови яичниково-маточных сосудов в окружающие ткани можно интерпретировать повышение активности лимфоцитов, моноцитов и эозинофилов и уменьшение активности нейтрофилов при минимальном лимфоцито-нейтрофильном отношении в позднюю лютеиновую фазу цикла у коров.
В крови магистральных сосудов, вдали от гормональных источников, распределение лейкоцитов было иное, закономерность которого проявляется в изменении лимфоцито-нейтрофильного отношения в различные периоды полового цикла. Минимальное лимфоцито-нейтрофильное от-ношение в фолликулиновой фазе цикла в крови из яремной вены соответствует минимальному лимфоцито-нейтрофильно-му отношению поздней лютеиновой фазе цикла в крови репродуктивных органов у коров. Обнаруженные различия, вероятно, обусловлены изменениями реактивности определенных групп лейкоцитов в усло-виях близости гормональных источников и особенностью яичниково-маточного кровотока.
Поддержание состояния физиологического лейкоцитоза в яичниково-маточных сосудах, по-видимому, способствовало ускоренному (оптимальному) использова-нию функционально разграниченных клеток белой крови необходимых для формирования структуры и функционирования репродуктивных органов коров в процессе их подготовки к оплодотворению и плодоношению.
Список литературы
1. Борисенков М.Ф., Монгалёв Н.П. Сравнительный анализ функции репродуктивных органов коровы и самки северного оленя: П. Клеточный состав крови в сосудах репродуктивных органов // Журн. эвол. биохим. и физиол. 2006. Т. 42. № 3. С.253-256.
2. Буянов А.А., Степанов Г.С., Седова Е.В., Савченко О.Н., Борисенков М.Ф. Морфофункциональные тесты послеубойного определения периода полового цикла коров // С- х. биол. 1991. № 2. С. 187-190.
3. Долгушин И.И., Бухарин О.В. Нейтрофилы и гомеостаз. Екатернбург: УрО РАН, 2001. 280 с.
4. Нежданов А.Г., Соловьев Н.А. Половые стероиды в крови коров при гипофункции яичников // Ветеринария, 1988. № 5. С. 41-43.
5. Соколовская И.И., Милованов В.К. Иммунология воспроизведения животных. М.: Колос, 1981. С. 264.
6. Сотникова Н.Ю., Воронин Д.Н. Особенности дифференцировки цитотоксических Т-лимфоцитов на системном и локальном уровне в динамике ге-стационного процесса // Вестник медицинской академической науки. Екатеринбург, 2010. № 2/1. С. 205.
7. Donald P. McDonnel. Molecular pharmacology of estrogen and progesterone receptors // Menopause Biology and Pathobiology. San Diego-Tokyo: Academic press, 2000. P. 3-11.
8. Ford S.P., Chenault J.R. Blood flow to the corpus luteum - bearing ovary and ipsilateral uterine horn of cows during the oestrous cycle and early pregnancy // J. Reprod Fertil. 1981. Vol. 62. № 2. P. 555-562.
9. Me Pherson L.A., Murdoch W.J. Localization of stress protein-70 in ovine corpora lutea during prostaglandin induced luteolysis // Prostaglandin. 1993. Vol. 46. № 5. P. 433-440.
10. Valle G.R., Castro A.C.S., Nogueira J.C., Caliari M.V, Graca D.S., Nascimento E.F. Eosinophils and mast cells in the oviduct of heifers under natural and superovulated estrous cycles // Anim. Reprod., 2009. Vol. 6. №2. P. 386-391.