Козин Р. В., Лебедев В. И., Иренкова Н. В.
Биология медоносной пчелы
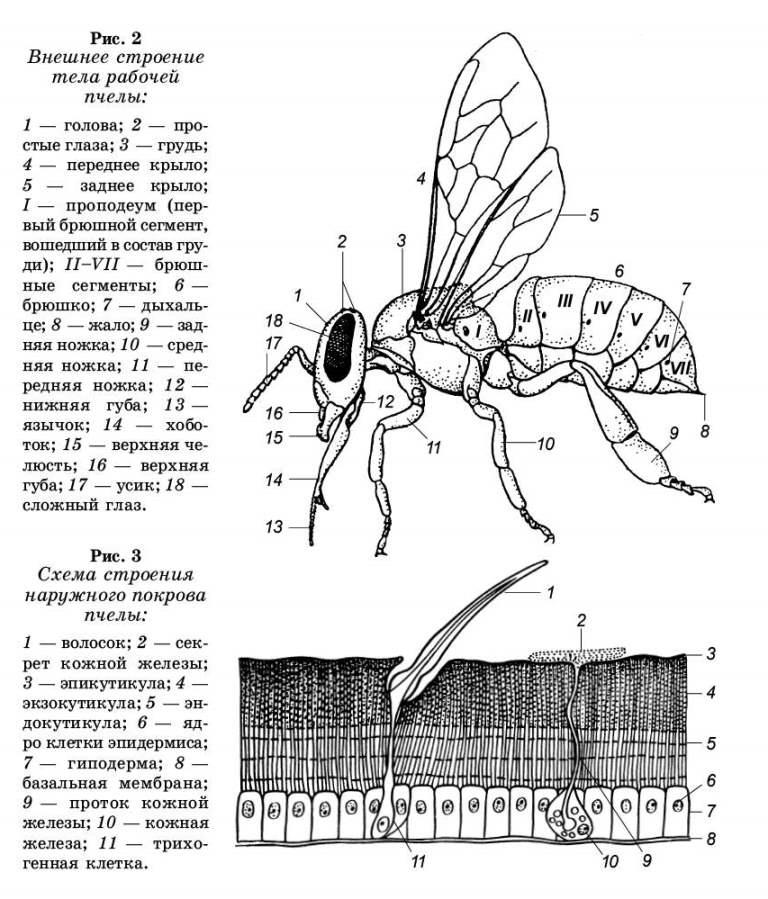
Тело медоносной пчелы, как и всех насекомых, разделяется на три подвижно соединенные между собой отдела: голову, грудь и брюшко (рис. 2).
В передней части от головы отходит пара усиков с многочисленными органами чувств, по бокам головы расположены два сложных (фасеточных) глаза, на темени три простых глаза. В нижней части головы расположена верхняя губа, по бокам ее находятся парные верхние челюсти и хоботок.
Внутри головы сосредоточена главная часть центральной нервной системы — головной мозг пчелы (надглоточный и подглоточный нервные узлы).
Грудной отдел пчелы состоит из четырех сросшихся между собой сегментов. От первых трех грудных сегментов отходят передняя, средняя и задняя пара ножек. Ко второму и третьему сегментам груди прикреплены также основания передней и задней пары крыльев. По бокам груди расположены три пары дыхалец — небольшие отверстия, через которые поступает воздух в трахейную систему. Внутри груди сосредоточены мощные мышцы, приводящие в дви-жение локомоторные органы — крылья и ножки.
Брюшко пчелы состоит из шести ясно видимых подвижных сегментов, на каждом из которых имеется пара дыхалец. Первый сегмент брюшка небольшой, на переднем конце сужается в стебелек, соединяющий брюшко с грудью. В конце брюшка находится жало, которое в спокойном состоянии пчелы скрыто внутри последних сегментов. Внутри брюшка пчелы сосредоточена основная часть внутренних органов пищеварения, дыхания, кровообращения, размножения и большая часть брюшной нервной цепочки.
Наружные покровы. Покровы насекомых состоят из двух разнородных образований — живых клеток эпидермиса или гиподермы и мертвой неклеточной кутикулы — продукта выделения этих клеток (рис. 3). Аморфная базальная мембрана подстилает гиподерму снизу, поддерживая ее клетки и ограничивая от гемолимфы. Она имеет мезодермальное происхождение и поэтому не относится к покровным тканям.
Эпидермис представлен непрерывным рядом столбчатых или многогранных клеток, снабженных крупными ядрами. На поверхности каждой клетки, примыкающей к кутикуле, имеются микроворсинки. Одна из самых важных функций эпидермиса — это секреторная. Она отчетливо проявляется в растворении старой кутикулы и продуцировании новых кутикулярных слоев во время линьки.
Кроме обычных неспециализированных клеток в эпидермисе насекомых есть специализированные клетки, образующие волоски и сенсиллы.
Кутикула образует наружный скелет, покрывающий все тело пчелы, служит опорой для прикрепления мышц и, следовательно, выполняет механическую функцию. Кутикула защищает тело от различных воздействий внешней среды. В своей основе кутикула эластична и гибка, но очень часто она подвергается уплотнению, или склероти-зации. Кутикула имеет сложное гистологическое строение. Она состоит из двух основных слоев: наружного — экзокутикулы и внутреннего — эндокутикулы. С внешней стороны экзокутикула часто бывает покрыта тонким липоидным слоем — эпикутикулой.
Эндокутикула — непосредственно прилегает к клеткам эпидермиса. Она прозрачна, состоит из множества тонких волокнистых пластинок, которые могут ограниченно скользить одна вдоль другой, что наряду с прозрачностью обеспечивает ее эластичность. В этой области процессы затвердевания и пигментации не выражены.
Экзокутикула — может рассматриваться как затвердевшая, окрашенная эндокутикулой, утратившая свое первоначальное пластинчатое строение. В состав экзокутикулы входят пигменты, окрашивающие хитин. Желтую, темную и коричневую окраску разных оттенков придает широко распространенный среди насекомых пигмент меланин. Доказано, что на окраску кутикулы влияет интенсивность освещения, температура и влажность, при которых происходит их развитие. Например, чем выше температура (в пре-делах от 16 до 35°С), тем светлее бывает окраска кутикулы. Поэтому местные пчелы на севере темного цвета, на юге их брюшко содержит светло-желтые полосы.
Все слои кутикулы снизу доверху пронизаны многочисленными покровными канальцами, которые начинаются от эпидермиса и обеспечивают связь с наружной поверхностью покровов. Внутри покровных канальцев проходят нитевидные отростки эпидермальных клеток.
Покровные канальцы участвуют в синтезе и транспортировании веществ, формирующих эпикутикулу. По канальцам транспортируется воск, дубильные вещества и материалы для построения кутикулы.
Эпикутикула — наружный, самый тонкий слой кутикулы, она не содержит хитина. Ее главную часть составляет кутикулин — вещество, включающее жиро- и воскопо-добные соединения. Восковой слой играет роль барьера, ограничивающего транспирацию и защищающего насекомых от потери воды. Доказано, что «критическая температура», при которой водопроницаемость покровов резко возрастает, совпадает с температурой плавления воскового слоя.
Воск эпикутикулы обычно покрывается сверху тон-ким цементным слоем, который выделяется многочис-ленными кожными железами. В состав этого слоя входят вещества, сходные с шеллаком. Они создают лаковое покрытие эпикутикулы и обеспечивают механическую защиту легко повреждаемого воскового слоя.
Химический состав кутикулы. Кутикула в основном состоит из органических веществ, а неорганические соединения составляют менее 1% от ее сухой массы. Наиболее важными органическими соединениями кутикулы являются хитин, белки, липиды и фенолы.
Хитин составляет от 25 до 60% от всего сухого вещества кутикулы. Совместно с белками он формирует основную часть наружного скелета насекомых. Физико-химические свойства хитина достаточно детально изучены. Он нерастворим в воде, щелочи и органических растворителях, но растворяется в крепких минеральных кислотах. В концентрированных растворах солей он растворяется, превращаясь в коллоидную массу. При нагревании с едким калием образуется хитозан, легко растворимое соединение в сильно разбавленных кислотах.
По химической природе хитин — высокомолекулярный азотосодержащий полисахарид, который напоминает гликоген или целлюлозу, но в отличии от этих соединений содержит аминогруппу NH, связанную с ацетильной группой О=С-СН3. Исходным продуктом для синтеза хитина служит глюкоза.
Белки составляют от одной четверти до половины сухого материала кутикулы насекомых. Прочность кутикуле насекомых придают белки, тесно связанные с хитином.
Липиды представлены преимущественно в восковом слое эпикутикулы. В составе кутикулярных липидов обнаружены свободные жирные кислоты, эфиры жирных кислот, углеводороды, диоловые спирты.
Вопрос о проницаемости кутикулы пчел играет огромное значение для понимания взаимоотношения их со средой и защиты от отравлений. Кутикула может не только затруднять, но и облегчать проникновение химических веществ через покровы. Высокая проницаемость покровов пчел для контактных инсектицидов связана со способностью этих ядов растворяться в восках эпикутикулы и с их абсорбцией хитином эндо- и экзокутикулы.
Некоторые инсектициды вызывают нарушение пассивного и активного транспорта воды через покровы. Так, при отравлении хлорорганическими инсектицидами из организма насекомых быстро уходит вода в результате испарения через кутикулу и дыхальца. К числу инсектицидов, в наибольшей степени нарушающих процессы водообмена, относятся фосфорорганические яды и соединения из группы пиретрина.
Установлено, что те яды, которые растворимы в органических растворителях, легко проникают через кутикулу в организм при контакте насекомого с ядом. Объясняется это присутствием в кутикуле липоидов, которые не являются для таких ядов барьером.
Поверхность кутикулы пчелы густо покрыта волосками разнообразной величины и строения, что, несомненно, играет определенную роль в регуляции температуры тела. Установлена прямая корреляционная зависимость между опушенностью тела пчелы и ее зимостойкостью. Волоски задерживают механические примеси воздуха при дыхании. На груди и ножках у пчелы развиваются разветвленные и пористые волоски, которые удерживают пыльцевые зерна цветков. На некоторых местах тела вырастают особо большие и прочные волоски-щетинки, выполняющие определенные механические функции. Начало волоску дает трихогенная клетка, расположенная в эпидермисе. Она образуется в процессе развития кутикулы.
Головной отдел. Голова пчелы (рис. 4) имеет треугольную форму и представляет собой сплошную, прочную, хитинизированную капсулу, не разделенную на членики. Она сплюснута спереди назад, а рот и челюсти обращены книзу. Голова расположена перпендикулярно продольной оси тела (гипогнатическии тип постановки головы).
Голова насекомого прошла сложный путь эволюции. В состав головы включались сегменты тела — шел процесс цефализации. Цефализация обусловлена тем, что передний конец тела, несущий ротовое отверстие и органы захватывания пищи (челюсти и др.), первым вступает в контакт с новым объектом среды. Поэтому в нем концентрируются органы чувств, а также передние отделы центральной нервной системы, регулирующие функционирование этих органов и составляющие головной мозг.
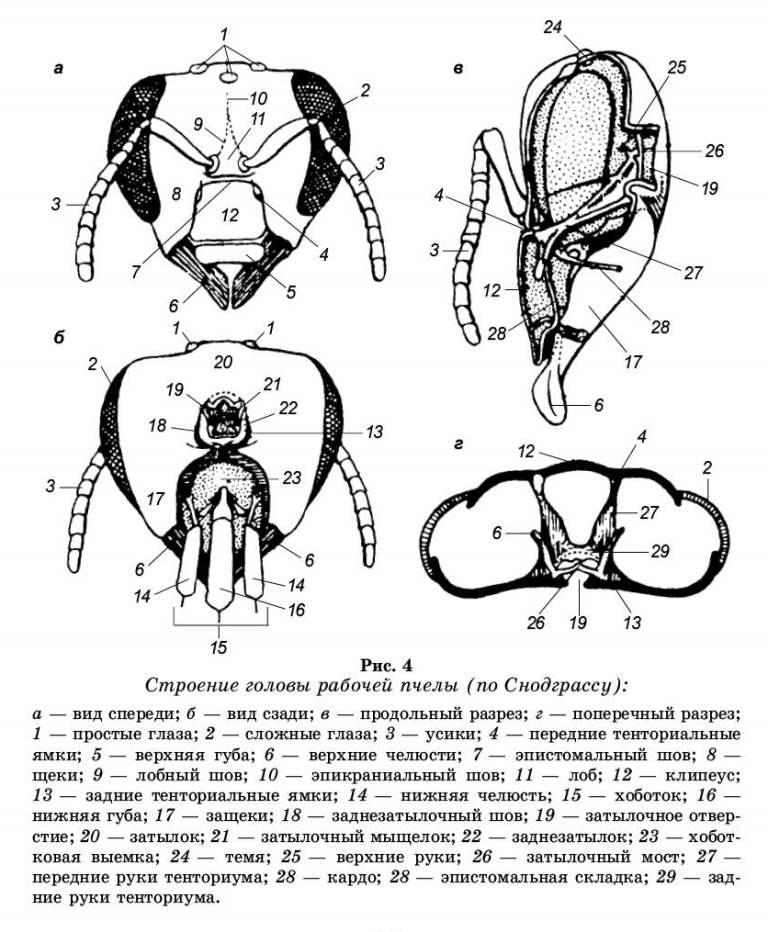
На головной капсуле различают поверхности, отделенные друг от друга швами. Передняя поверхность называется лицевой, верхняя — теменем, задняя — затылком. Вдоль темени посередине головы проходит эпикраниальный шов, который делит темя на правую и левую половины. На темени по обе стороны от шва расположены два простых глаза, третий глаз размещен несколько ниже, на линии эпикрани-ального шва. По бокам головы расположены два сложных (фасеточных) глаза. Ниже, на лицевой стороне, эпикраниальный шов делится на две ветки и образует треугольное поле — лоб. От основания лба отходит пара подвижных членистых усиков (антенны). Усик состоит из трех основных частей: основного членика (скапуса), маленького педи-целлярного членика (ножки) и длинного и очень подвижного жгутика. Первый членик на внутреннем конце имеет расширение в виде круглой головки, которая входит в мягкую сочленованную ямку, окруженную валиком. Скапус направлен косо к внешней плоскости головы. Внутри этого членика имеются мышцы, обеспечивающие вращение усиков во все стороны. К основному членику прикреплен второй маленький членик — ножка. Почти у всех насекомых на этом членике находится орган равновесия (джонстонов орган). Ножка может несколько перемещаться относительно первого членика за счет мышц, соединяющих ножку и скапус. За ножкой следует жгутик, состоящий из члеников очень близких по строению. Задний конец каждого членика немного сужается и входит в слегка расширенный передний конец последующего членика. Жгутик не имеет мышц, но обладает небольшой подвижностью вследствие членистого строения и особого сочленения. Жгутик матки и рабочих пчел состоит из 10 члеников, а жгутик трутня из 11 члеников, каждый из которых больше по величине.
На члениках жгутика, начиная с третьего (два первых членика меньшего размера, чем последующие), размещены многочисленные чувствительные органы — сенсиллы. У отдельных насекомых насчитываются десятки тысяч сенсилл различного назначения. Например, у медоносной пчелы 14-15 тыс. обонятельных ямок на каждом усике, а у самца майского жука около 50 тыс., у самки — до 8 тыс.
В нижней части лоб ограничен горизонтальным эпи-стомальным швом. Ниже этого шва размещена прямоугольная пластинка — лицевой щиток (клипеус). К вому щитку прикреплена верхняя губа. По бокам лицевого щитка (по обе стороны от лобных швов) расположены щеки, которые загибаются на бока головы. По бокам головы щеки граничат у всех особей со сложными глазами. К нижней части щек прикреплены верхние челюсти (мандибулы).
В затылочной части головы имеется затылочное отверстие, через которое голова сообщается с грудью. Ниже затылочного отверстия расположена хоботковая выемка, в которой закреплены основания нижних челюстей и нижней губы. В хоботковую выемку складывается хоботок, будучи в нерабочем положении.
На заднезатылке расположены два затылочных мы-щелка, обеспечивающие сочленение головы к груди.
Прочность головы усиливается внутренним скелетом (тен-ториумом), расположенным в виде хитиновых перемычек между передней и задней стенками головы. От верхних углов клипеуса (внутри головы) назад и несколько вверх отходят передние руки тенториума. От затылочного отверстия в глубину головы идут задние руки, которые направляются вперед и вниз к середине, где сливаются с передними руками тенториума. От передних рук вверх отходит пара верхних рук, которые заканчиваются свободно в виде выростов. Около затылочного отверстия задние руки тенториума соединяются перемычкой, образующей затылочный мост.
Внутренний скелет головы придает необходимую устойчивость, жесткость в нижней части головной капсулы, где размещены ротовые органы. Увеличение прочности головы необходимо в связи со следующим: ротовым аппаратом пчелы собирают пищу (жидкую и твердую), размягчают воск и грызут дерево при расширении летка; головой утрамбовывают пыльцу в ячейках сота. Одновременно внутренний скелет обеспечивает защиту от повреждения головного мозга и других органов (глотки, слюнных желез, трахей, кон-цевой части аорты), расположенных внутри головы. Он же служит опорой для глотки и местом прикрепления мышц.
Ротовой аппарат насекомых в процессе эволюции прошел сложный путь развития. По мере того как в головной отдел втягивались сегменты туловища, ротовое отверстие окружалось конечностями втянутых туловищных сегментов. Постепенно эти конечности утрачивали свою локомоторную функцию и приобретали новую, связанную с захватом и измельчением пищи. Ротовой аппарат насекомых имеет много типов и форм строения в зависимости от образа жизни насекомого и способа добычи пищи (грызущие, лижущие, колющие, сосущие и др.)- Исходной (первичной) формой у насекомых является грызущий тип ротового аппарата, а от него произошли все другие типы или модификации. Типичный грызуще-жующий тип ротового аппарата имеют примитивные группы насекомых (таракановые). Такой ротовой аппарат приспособлен для приема твердой пищи, в связи с чем все его органы укорочены и сильно хитинизированы.
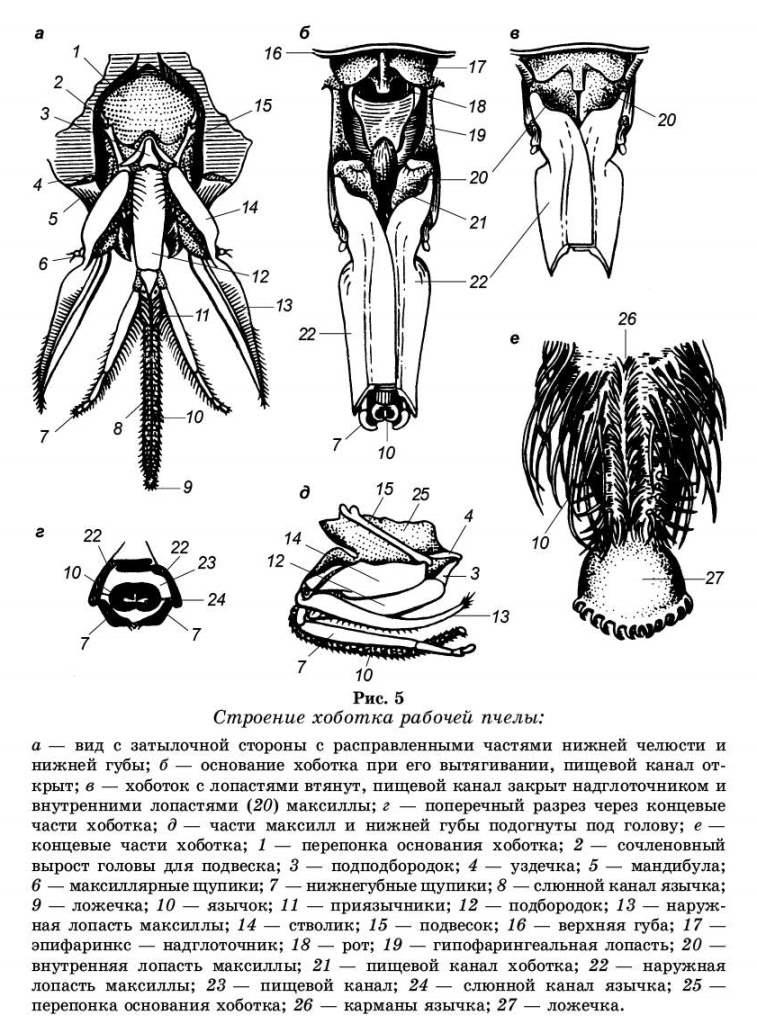
Ротовой аппарат пчелы относится к грызуще-сосуще-лижущему типу. Ротовой аппарат пчелы представлен двумя независимыми органами, образующими грызущую и сосущую системы. К грызущей системе относятся мандибулы и верхняя губа, к сосущей — максилла (нижняя челюсть) и нижняя губа. Нижние челюсти совместно с нижней губой образовали вытянутый хоботок, которым пчела засасывает жидкую пищу (рис. 5).
Верхняя губа — это подвижная сильно склеротизиро-ванная пластинка, прикрывающая вход в ротовую полость. Она свободно свисает с нижнего края лицевого щитка. Под верхней губой и ротовым отверстием находится мягкий выступ — эпифаринкс. Когда части хоботка складываются в трубку, между основными члениками нижних челюстей образуется щель. Ее и прикрывает сверху эпифаринкс. К эпифаринксу подходят мышцы от клипеуса, играющие определенную роль в процессе заглатывания пищи.
Внутренняя стенка, общая для клипеуса и верхней губы, образует переднюю стенку расширенной полости — цибариума. Эта полость расположена сразу же за ротовым отверстием. Между клипеусом и передней стенкой цибариума проходят пять коротких мышц, сокращение которых приводит к увеличению полости цибариума.
Верхние челюсти — мандибулы, называемые также жвалами, укороченные, твердые, нерасчлененные хити-новые образования, суженные в середине, вогнутые к внутренней стороне. Они прикреплены по бокам верхней губы и поддерживают хоботок в свернутом и развернутом состоянии. Мандибулы своим основанием причленя-ются к голове таким образом, что могут перемещаться в поперечном направлении. К наружной стороне мандибулы прикреплена мышца, сокращение которой отводит ее в сторону. Вторая, более мощная, приводящая мышца прикреплена к внутренней стороне мандибулы. Попеременное сокращение этих мышц сводит и разводит мандибулы в стороны. Внутри мандибулы размещена верхнечелюстная железа, выводной проток которой выходит наружу.
Форма мандибул у всех стаз пчелиной семьи различная. У рабочей пчелы они наиболее сильно и хорошо развиты.
Жевательные края их закруглены, на верхней стороне имеется расширение и углубление в виде ложки. Здесь же располагаются небольшие гребневидные валики.
Верхними челюстями рабочие пчелы выполняют разнообразные работы: разгрызают пыльники на тычинках цветов при сборе пыльцы, откусывают кусочки перги в ячейках, разминают воск при строительстве сотов, разгрызают дерево при расширении очень узкого летка, прогрызают крышечки ячеек по окончании метаморфоза, удаляют крышечки с ячеек сота с запечатанным медом. Кроме того, рабочие пчелы мандибулы используют при охране гнезда, хватая ими чужих пчел, врагов и вредителей.
У матки мандибулы более широкие и на конце имеют острый зубец, которым она надрезает восковую крышечку перед выходом из маточника. Мандибулы трутня недоразвиты. Жевательный край сглажен, имеется небольшой зубец, которым он разрушает крышечку перед выходом из ячейки. Вся челюсть покрыта волосками.
Нижняя челюсть (максилла) парная и состоит из основного членика (кардо) или подвеска, стволика (стипеса) и наружной саблевидной лопасти, или галеа. Подвеска подвижно соединена посредством мыщелка со впадиной на краю головной капсулы. Стволик причленен к переднему концу подвеска. На конце стволика снаружи находится рудиментарный максиллярный щупик. В вытянутом положении галеа правой и левой нижних челюстей складываются вместе, соединяясь вдоль ребра. Передние части плотно накладываются одна на другую, образуя переднюю и боковые стороны канала хоботка.
К стволику прикреплены три грудные мышцы (при сокращении которых хоботок выдвигается вперед в рабочее положение во время всасывания пищи) и внутренняя жевательная лопасть (которая служит для регулирования функционирования пищевого и секреторного каналов). За счет перемещения жевательных лопастей образуется прямой канал во время засасывания нектара хоботком. Во время кормления молодых личинок рабочих пчел или матки жевательные лопасти оттягиваются, открывая тем самым предротовую полость и обеспечивая выход секрета глоточных желез наружу. При таком по-ложении ротовых органов пчела уже не может засасы-вать хоботком пищу.
Нижняя губа — устроена наиболее сложно. У пчелы в нижней губе сохранились все типичные для насекомых части, но они сильно видоизменены. В нижней губе различают под подбородок (основание нижней губы), имеющий форму небольшого треугольника, и второй удлиненный, хорошо хитинизированный членик — подбородок. Вниз от подбородка отходит сильно вытянутый язычок, оканчивающийся слегка расширенной ложечкой. По бокам от основания язычка к подбородку прикрепляются два приязыч-ника (параглоссы) и два четырехчлениковых щупика.
Нижние челюсти и нижняя губа объединяются при помощи изогнутой пластинки — уздечки и хитиновой мембраны, натянутой между подвесками.
Щупики пчелы утратили функцию органов чувств, а входят в составную часть хоботка — органа приема пищи. Во время приема пищи пчелой щупики образуют заднюю и частично боковые стенки большой трубки хоботка. Параглоссы охватывают основание язычка с обеих сторон в виде коротких полукруглых пластинок, способных отгибаться и плотно прижимать язычок. В прижатом положении они обеспечивают проход секрета слюнных желез в канал язычка, а в раздвинутом — открывают прямой проход пищи через большую трубку в рот. Язычок пчелы состоит из чередующихся колец мягкой эластичной и плотной кутикулы. Кольца плотной кутикулы несут на себе волоски, обращенные к концу язычка. Такое строение придает язычку гибкость, прочность неэластичность. Кольца, образующие язычок, в одном месте не смыкаются, за счет чего по всей длине язычка образуется тонкий капиллярный канал.
Таким образом, хоботок пчелы образует три канала разного диаметра. Самый маленький (капиллярный) ка-нал проходит внутри осевого стержня язычка. По капиллярному каналу к концу хоботка поступает секрет слюнных желез. Канал среднего размера, образуемый внешней кутикулой, служит для прохода жидкой пищи в тех случаях, когда пчела слизывает мельчайшие капельки нектара ложечкой. Третий канал наибольшего диаметра, образуемый при сложении вместе галеа максиллы и щупиков нижней губы, служит для всасывания нектара, когда его в цветке много. При этом способе взятия корма пчела погружает хоботок в каплю наполовину его длины. Внутри этой трубки язычок во время всасывания совершает быстрые движения вперед и назад (подобно поршню насоса), что вызывает перемещение нектара к основанию язычка. Передвижению нектара в медовый зобик способствуют наса-сывательные движения глотки — глоточный насос, мышцы которого натянуты между глоткой и лицевым щитком.
Строение хоботка у всех особей пчелиной семьи одинаковое. Разница заключается только лишь в его длине. Длина хоботков у маток и трутней различных пород пчел почти одинакова. У маток среднерусских пчел она равна 4,14 мм, у трутней — 4,04 мм, итальянских — 4,06 и 4,08 мм, краинских — 4,24 и 4,48 мм, и кавказских — 4,35 и 4,23 мм, соответственно. Наиболее длинный хоботок у рабочих пчел, но его длина колеблется в больших пределах, чем у маток и трутней, в зависимости от принадлеж-ности к той или иной породе, от 5,5 до 7,2 мм. Величина хоботка устойчиво передается по наследству. Этот признак незаменим при определении породной принадлежности пчел, а также имеет самостоятельное селекционное, биологическое и хозяйственное значение: пчелы, обладающие длинным хоботком, способны доставать нектар из нектарников, расположенных глубоко в цветках. В связи с этим в практической работе по бонитировке пчелиных семей, селекционной и научной работе измеряют длину хоботка. За длину хоботка принимают расстояние от вершины подбородка до наружной границы ложечки нижней губы. Величина хоботка в значительной мере подвержена сезонным изменениям, что необходимо учитывать во избежание ошибок.
Грудной отдел медоносной пчелы имеет усложненное строение по сравнению с примитивными насекомыми. Поэтому целесообразно первоначально ознакомиться с более простой схемой строения грудного отдела и его сегментов. Грудь насекомого состоит из трех кутикулярных члеников, или сегментов переднегруди, среднегруди, заднегру-ди. Каждый такой сегмент подразделяется на четыре отдельных склерита: спинное полукольцо — тергит, брюшное полукольцо — стерпит и пара первоначально мягких боковых стенок — плейритов.
У примитивных отрядов насекомых членики грудного отдела хорошо обособлены друг от друга и более или менее равномерно развиты. Вследствие того, что на груди у насекомых сосредоточены органы передвижения, мускулатура грудных сегментов усиливалась и усложнялась, увеличивались размеры сегментов, особенно их диаметр, ус-ложнялся наружный скелет.
В состав груди пчелы, как и других хорошо летающих насекомых, входят четыре сегмента: три собственно грудные, четвертый (перешедшего из брюшка), сросшийся по всей площади с заднегрудным сегментом (рис. 6). Первый сегмент груди называется — переднегрудь, второй — сред-негрудь, третий — заднегрудь и четвертый — проподеум. Грудь пчелы отделена от брюшка перетяжкой (стебелек), образуемой между первым и вторым брюшным сегментами. Первый грудной сегмент у медоносных пчел преобразовался в связи с причленением к голове, второй сильно разросся, а третий уменьшился до узкого кольца.
Тергит переднегруди представляет собой узкую подковообразную пластинку. Тергит прочно срастается с грудной капсулой, а плейриты и стернит, имеющий форму треугольной пластинки с углом, обращенным вперед, соединяются с головой, образуя подобие шеи.
Среднегрудь пчелы составляет основную часть груди. Тергит среднегруди делится на скутум и скутеллюм. Плейриты сильно разрослись и соединились со стернитом. Посередине стернита проходит стернальный гребень. По бокам скутума отходят два передних отростка, а по бокам скутеллюма — задние крыловые отростки, служащие для присоединения двух пар крыльев.
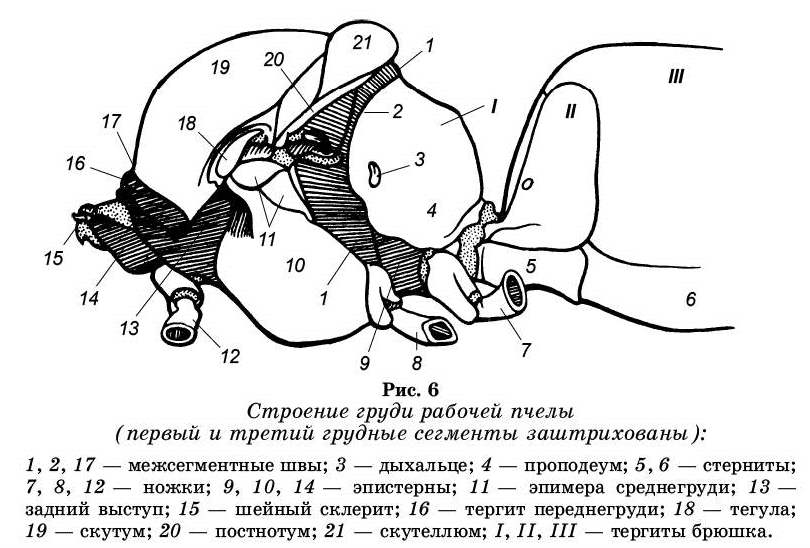
Заднегрудь представлена узким кольцом. Тергит не имеет подразделений, а плейриты косой бороздкой делятся на две части.
Проподеум состоит из широкого, сильно выпуклого тергита и стернита, представленного узкой полоской.
Переход первого членика брюшка в состав груди обусловлен необходимостью увеличения объема этого отдела, служащего вместилищем для сильно развитой мускулатуры, обеспечивающей полет пчелы. Развитие мощной мускулатуры в грудном отделе пчелы одновременно повело за собой сильное развитие и усложненность внутреннего скелета.
Внутренний скелет (тенториум) представлен различными хитиновыми перегородками, и выростами, вдающимися в полость тела (см. рис. 12). К внутреннему скелету прикрепляются мышцы, обеспечивающие полет пчелы.
На границе передне- и среднегруди находится хитиновый гребень — первая грудная фрагма. Между скутумом и скутеллюмом имеется вторая фрагма (мезофрагма). Она образует неправильной формы гребень. От него отходит большой лопатообразный отросток назад, который далеко заходит за пределы тергита заднегруди, вдаваясь в полость про-подеума. От стернального гребня внутрь идет большой хитиновый гребень — грудная кость (эндостернум).
На груди пчелы расположены три пары дыхалец: первая пара находится на границе передне- и среднегруди под выростом тергита переднегруди; вторая — на заднегруди под основанием задних крыльев (дыхальце очень маленькое и имеет вид щели), третья (самые крупные дыхальца, овальной формы) — на тергите проподеума.
Грудь является локомоторным органом, на ней находятся органы передвижения — две пары крыльев и три пары ножек.
Локомоторные органы. У медоносной пчелы три пары ножек, которые состоят из следующих частей: тазик или кокса, вертлуг или троксантер, бедро или фемур, голень или тибиа, лапки или тарзус и коготковый членик или претарзус (рис. 7).
Тазик (кокса) — это отдел ножки, сочлененный с телом; в месте сочленения тазика с грудью находится впадина. Тазик вместе с плейритом образует туловищное соединение, которое позволяет конечности двигаться вперед и назад.
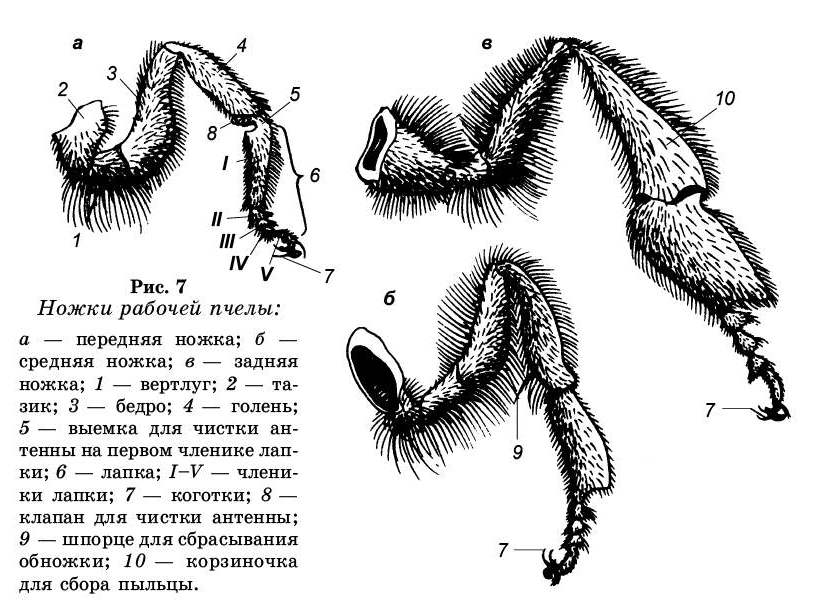
Вертлуг (троксантер) — маленький членик, который подвижно соединен с тазиком сочленением с вращательным суставом, обеспечивающим передвижение ноги вверх и вниз, а также до некоторой степени вперед и назад и малоподвижно с бедром.
Бедро (фемур) — круглая часть ножки. Сочленение бедра с голенью (называется коленным или тибио-фермуаль-ным) дает возможность ножке распрямляться, но не позволяет ей перегибаться в обратную сторону.
Голень (тибиа) — приблизительно равна по длине бедру, но различается по ширине на передних, средних и задних ножках.
Лапка (тарзус) — в отличие от всех предшествующих частей состоит из члеников: одного большого и четырех маленьких.
Движение ножек обеспечивается двумя группами мышц, одна из которых, начинаясь в груди, заканчивается в ножке, другая целиком лежит в пределах ножки.
Первая группа состоит из трех мышц, которые начинаются на фурке и идут — первая к переднему краю тазика, вторая — к вертлужному сухожилию, третья — к заднему краю тазика. Первая мышца приводит, а третья отводит тазик. Вторая мышца вместе с пятой дорсально-ветральной является опускателем вертлуга и бедра.
В тазике находятся две мышцы. Из них первая идет от внешней стенки тазика к наружному краю вертлуга, вторая, начинаясь от внутренней части тазика, присоединяется к вертлужному сухожилию. Первая отводит, вторая сгибает вертлуг и бедро.
В вертлуге имеется только одна мышца, которая вызывает вращение бедра.
Движение голени осуществляется двумя мышцами, которые целиком лежат в бедре. Кроме того, в бедре находится третья мышца, от которой отходит длинное претарзальное сухожилие, переходящее в голень и продолжающееся там.
В голени располагаются три мышцы, сгибающие и выпрямляющие лапку. Внутри лапки мышц нет.
Ножки служат пчеле для различных целей: передви-жения, опоры тела, сбора и переноса пыльцы, чистки усиков и тела.
Передвижения пчелы по шероховатой и по гладкой поверхности обеспечивает коготковый членик (претарзус) и по-
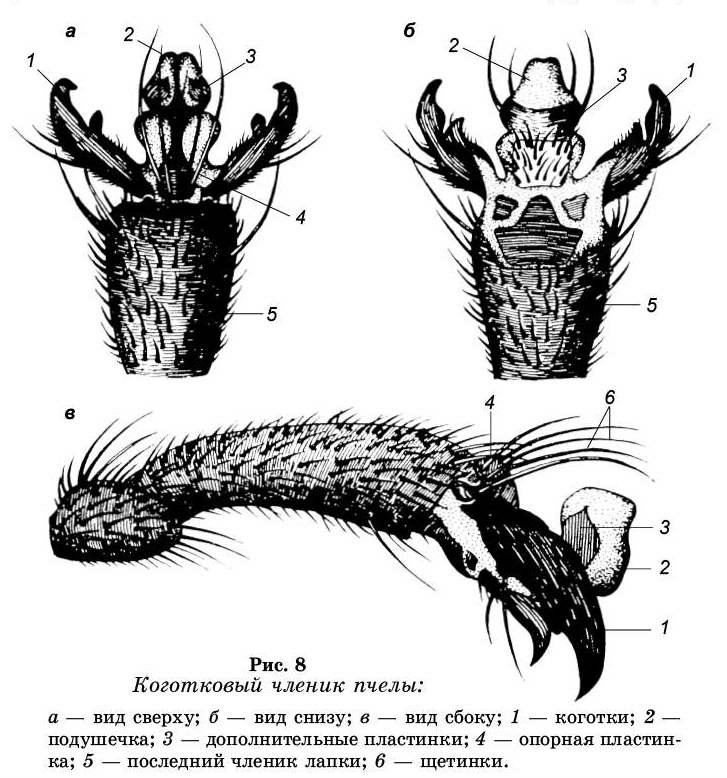
душечка, расположенные на лапке (рис. 8). Коготковый членик имеет два коготка, они сильно раздвоены и концы их загнуты вниз. Между коготками расположена эластичная подушечка. Над подушечкой находится пять длинных загнутых щетинок, которые предохраняют ее тонкие стенки. Подушечка представляет собой мешочек, и когда пчела ходит по гладкой поверхности, то она с их помощью приса-сывается, к ней. Присасывание к поверхности обеспечивается за счет того, что под подушечкой создается вакуум. Не исключено, что главную роль в присасывании подушечки играют клейкие выделения на нижней ее поверхности.
Несмотря на то, что все три пары ножек устроены в основном одинаково, в деталях они между собой различаются в соответствии с выполняемыми функциями.
Передние ножки. Они меньше других и наиболее подвижны. На внутреннем крае первого членика лапки передней ножки имеется полулунная выемка, усаженная хитиновыми щетинками. Против выемки находится отросток-клапан с дополнительной пластинкой, отходящей от заднего конца голени. Вкладывая усик (антенну) в выемку и прижимая его отростком, пчела протягивает усик назад и вперед. При этом гребнем выемки очищается его наружная поверхность, а посторонние частицы с внутренней стороны соскабливаются дополнительно пластинкой клапана. Аппарат для чистки усиков — важное биологическое приспособление, так как на поверхности усиков сосредоточены многочисленные органы чувств. Аппарат для чистки усиков имеется у всех особей пчелиной семьи. Кроме того, первый членик лапки покрыт длинными волосками, образующими пыльцевую щеточку, используемую для чистки ротовых частей, а также для сбора пыльцы с передней части тела.
Средние ножки. Они могут двигаться только назад и вперед по вертикальной оси. На внутренней стороне в нижней части голени имеется игловидный отросток, который называется шпорцем. Представляет он собой хитиновую палочку. Шпорце служит для складывания пыльцы в ячейку. Имеется только у рабочей пчелы.
Задние ножки. Они более подвижны, чем средние. С их помощью рабочая пчела приносит в улей пыльцу, пропо-лисное сырье. Голень задней ноги медоносной пчелы расширена, и ее наружная сторона имеет гладкую вогнутую поверхность, окаймленную жесткими длинными волосками. Желобок, тянущийся по всей поверхности голени, называется корзиночкой. В ней укладывается обножка (комочек пыльцы) при работе пчелы на цветках. К вершине голени корзиночка расширяется, несколько отступая от волосков; на две корзиночки имеется один большой волос, изогнутый к входу. По самому краю голени находится ряд острых длинных зубцов, образующих гребень. Расширенную форму имеет и первый членик лапки. С наружной стороны у него расположены 9-10 рядов тонких щетинок, расположенных под углом 45° к поверхности лапки, носящих название щеточки. Щеточка так же как и корзиночка необходима пчеле при сборе пыльцы. Кроме того, щеточками задних ножек пчелы извлекают восковые пластинки с восковых зеркалец брюшных полуколец. Первый членик лапки (пятка) причленен к голени не всем основанием, а только его передним углом. В результате этого членик может совершать маятникообразные движения. Пространство между голенью и пяткой называется щипчиками. Все опи-санные выше приспособления в первую очередь служат для сбора пыльцы, поэтому ни трутень, ни матка их не имеют.
Когда пчела работает на цветке, пыльца прилипает к многочисленным густым волоскам груди и голове. Кроме того, часть пыльцы пчела при помощи челюстей и передних лапок сцарапывает непосредственно с тычинок. Затем эта пыльца складывается на щеточках средних ног. После этого щеточки средних ножек зажимаются между щеточками задних, так что пыльца со средней правой ножки переходит на заднюю левую. Таким же образом с левой средней ноги пыльца попадает на заднюю правую. Задние ножки своими жесткими щеточками чистят также брюшко.
С головы, с придатков и первого сегмента грудного отдела пыльцу пчела счищает главным образом щеточками передних ножек. При этом она смачивает пыльцу некоторым количеством слюны и нектара. Щеточками средних ножек пчела счищает пыльцу с затылка последних двух сегментов груди.
Когда на щеточках набирается достаточное количество пыльцы, пчела под брюшком сближает задние ножки таким образом, что гребень одной ножки вычесывается щеточкой другой. В результате на наружной поверхности каждого гребня формируется по комочку пыльцы. Завершающий этап работы заключается в том, что лапка начинает двигаться вперед и назад. В результате этого комочек пыльцы сдвигается внутрь корзиночки, в середине которой находится одна крупная щетинка. При помощи щетинки и боковых волосков пыльца удерживается. Этот процесс повторяется многократно, и, в конце концов, в корзиночке образуется большой комочек пыльцы, называемый обножкой.
Когда пыльцевой комочек становится достаточно большим, пчела возвращается в улей, отыскивает ячейку, предназначенную для запасов корма и, упираясь средней ножкой в верхний конец комочка, выталкивает его из корзиночки.
Крыльев у пчелы две пары и являются они придатками средне- и заднегруди. Крылья развиваются полностью только при переходе пчелы из стадии куколки во взрослое состояние. Закладываются они в виде мешкообразного зачатка покровов средне- и заднегруди. С ростом куколки этот зачаток разрастается, его брюшная и спинная стенки плотно сближаются. У взрослой пчелиной особи крыло — тонкая, эластичная пластинка, по которой проходят жилки, представляющие собой затвердевшие участки полых трубок. По ним во время формирования крыла нагнетается гемолимфа, расправляя крыловую пластинку.
Жилки — механическая опора крыла, они помогают преодолеть сопротивление воздуха при полете. В крыле различаются, продольные жилки, частично ветвящиеся, и поперечные, соединяющие продольные друг с другом. Между жилками натянуты тонкие прозрачные перепонки. Рисунок из продольных и поперечных жилок называется жилкованием. Просветы перепончатой части крыла, заключенные между жилками, называются ячейками. Каждая жилка в зависимости от ее расположения на крыле имеет определенное название (см. рис. 9).
На крыльях пчелы от основания отходят пять про-дольных жилок: костальная, субкостальная, медиальная, анальная и невральная.
Костальная жилка, образующая утолщенный передний край крыла, отходит от плечевой пластинки, не ветвится.
Субкостальная жилка идет рядом с костальной, в середине крыла сливается с ней. Она также не ветвится.
Медиальная, или срединная жилка, короткая, разде-ляется на две ветви — базальную, соединяющуюся с субкостальной, и дискоидальную.
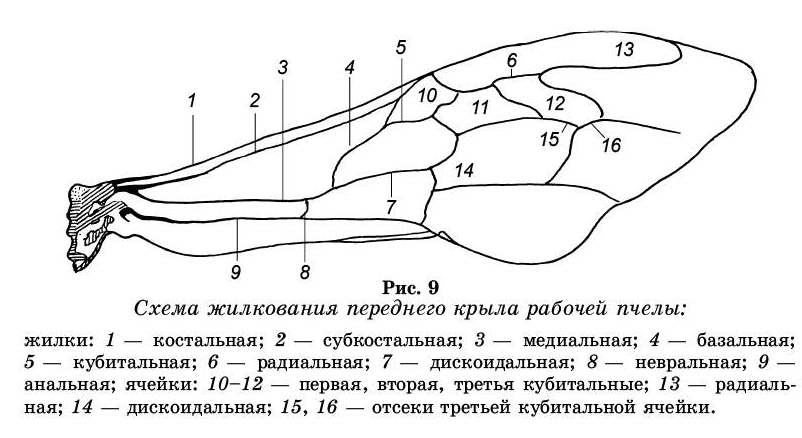
Кубитальная жилка отходит от середины жилки и тянется вдоль крыла почти до его конца.
Радиальная жилка связана с кубитальнои тремя меж-кубитальными жилками.
Анальная жилка сначала идет параллельно медиальной, затем расходится с ней. Между ними находится короткая невральная жилка.
На крыльях пчелы развиты также следующие про-дольные жилки: субдискоидальная, первая возвратная, вторая возвратная.
Жилки образуют на крыле замкнутые ячейки, кото-рые так же как и жилки имеют названия: радиальная, кубитальная, дискоидальная. Их названия определяются по прилегающим продольным жилкам.
При установлении породы медоносных пчел хорошо характеризует их породную принадлежность кубитальныи индекс, который определяется по формуле: а/б 100% , т. е. отношением длины одной жилки (а) третьей кубитальнои ячейки к другой (б). Этот признак практически не подвергается сезонным изменениям, слабо коррелирует с остальными экстерьерными признаками.
Заднее крыло имеет сходное строение, но значительно меньше переднего (в 1,5 раза короче). Кроме того, у трутней встречается добавочное жилкование задних крыльев. Специфика жилкования заднего крыла отражает индивидуальные особенности каждой матки, а отсюда и отдельных пчелиных семей. Следовательно, можно считать, что специфика жилкования задних крыльев является морфологическим признаком при определении принадлежности пчел к той или иной группе или линии одной породы.
Заднее крыло снабжено на переднем крае рядом крючков, направленных вверх. При движении пчелы переднее крыло с расположенной на заднем крае складкой скользит по заднему и крючки цепляются за нее. Этим достигается прочное сцепление передней пары с задней. При складывании они легко разъединяются. У рабочей пчелы на заднем крыле число крючков колеблется от 14 до 28; у матки — от 13 до 23; у трутней — от 13 до 29.
Наблюдаются также породные различия в количестве зацепок. Так, например, северные пчелы имеют меньше зацепок, чем южные.
Сочление крыла с телом представляет собой сложный механизм, который обеспечивает крылу быстроту взмаха и определенные повороты крыла при взмахе. Кроме того, сочление обеспечивает складывание крыла. Крыло прикреплено к мембране между спинкой и плейритами груди. Место прикрепления мышц к мембране подвижно.
Роль точки опоры для крыла играет столбик-вырост плейрита. Снаружи от столбика лежит длинное плечо; от места прикрепления к мембране сегмента до столбика — короткое плечо. Смещение спинки ведет к поднятию крыла. Качание крыла обеспечивается благодаря тому, что концы жилок не доходят до края крыла.
Существенную роль в работе крыльев играет серия со-членовых пластинок, несколько аксиллярных и промежуточная. Эти пластинки укрепляют основание крыла, обеспечивают его сгибание по наружным линиям и передачу движения к концам жилок.
Полет пчелы сильно отличается от полета птицы тем, что у пчелы крылья лишены мышц.
Более того, крылья насекомых только аналогичны крыльям позвоночных, но не гомологичны. В отличие от птиц у большинства насекомых мышцы прикреплены не к крыльям, а к стенке тела, причем таким образом, что даже самые слабые изменения формы грудного отдела заставляют крылья быстро подниматься и опускаться.
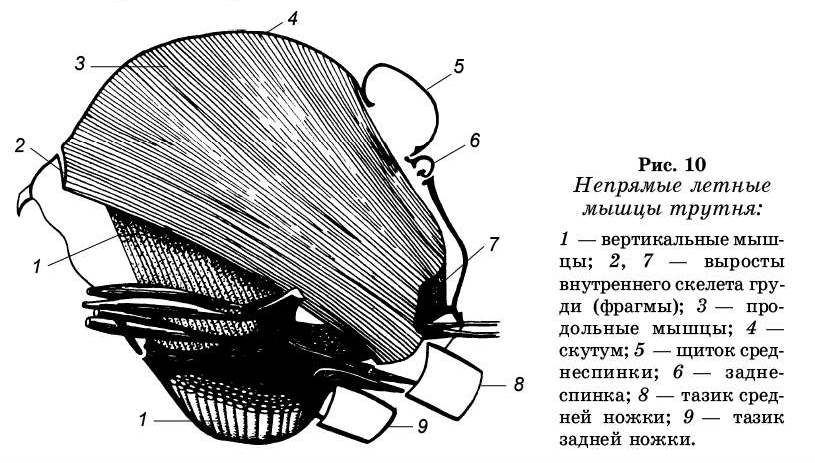
Движение крыльев в полете координируется и на-правляется в первую очередь энергией мышц непрямого действия, т. е. мышц, не соединенных с крылом. К этой группе относятся продольные спинные и вертикальные мышцы (рис. 10). Сокращение вертикальных мышц от-тягивает вниз тергит. В результате крылья на противоположной стороне от точки опоры поднимаются. При сокращении продольных мышц тергит выгибается, а крыло опускается. Движения стенки тела едва заметны, но, поскольку плечи рычага по обе стороны от точки опоры крыла имеют различную длину, размах движения кончиков крыльев в несколько сот раз больше. Вторая группа мышц — мышцы прямого действия, которые обеспечивают поворот крыла вдоль продольной оси, а также отводят его вперед и назад.
Основную роль при полете играют непрямые мышцы (рис. 11). Полость груди у медоносной пчелы целиком заполнена ими. Их две пары. Первая пара прикреплена к передней и задней фрагмам среднегруди. Сокращение этих мышц приводит к некоторому сближению фрагм в сегменте за счет изгиба вверх и увеличения ее кривизны. В результате основание крыла поднимается, а гребная часть крыла опускается. Вторая пара продольных спинных мышц значительно меньше первой. Натянута она между задней фрагмой и скутумом. Эти мышцы также участвуют в опускании крыла. Следовательно, мышцы продольной системы играют роль опускателей.
Другая группа мышц непрямого действия — верти-кальная мускулатура. Они идут от спинки груди к стерни-ту груди, при сокращении которых притягивается спинка к грудке, вследствие чего осуществляется подъем гребной части крыла вверх.
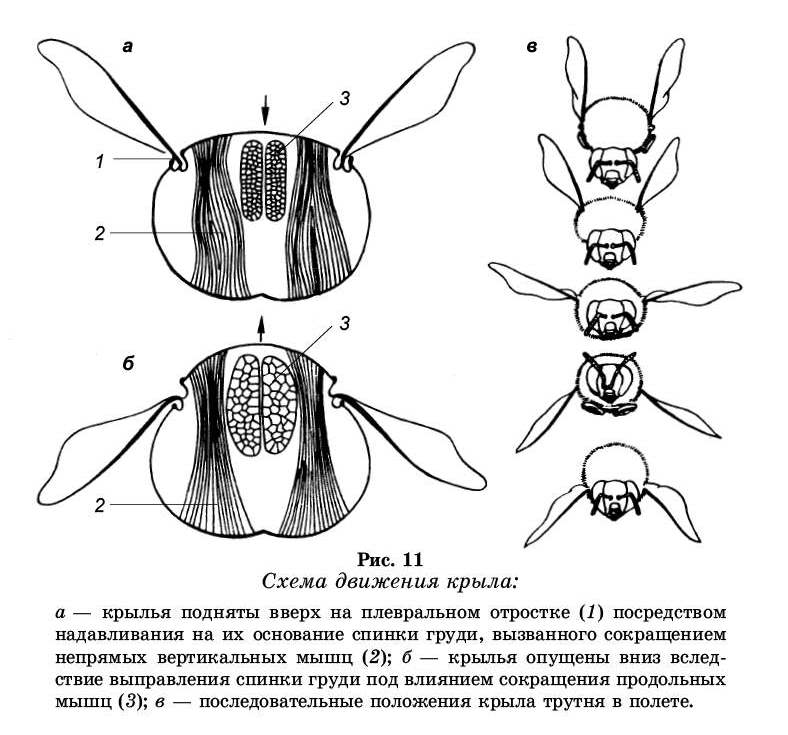
рода пропеллер. Движение крыла при полете характеризуется сложной траекторией, имеющей вид цифры 8, наклоненной верхним концом назад; при свободном движении вперед эта восьмерка растягивается и превращается в синусоиду. Сложные движения крыльев в полете являются комбинацией более простых движений вверх и вниз, вперед и назад, изменений наклона. При этом пчела делает большое количество вертикальных взмахов крыла (от 85 до 235 в секунду).
Для насекомых характерны три вида полетов: планирующий, парашютный и гребной. Для пчел присущ гребной полет, который является основной формой полета насекомых. Осуществляется он при помощи непрерывных ритмических ударов крыльями.
Скорость полета рабочей пчелы во многом зависит от погодных условий (сила ветра), расстояния, нагрузки. Средняя скорость полета пчел около 24 км/ч, максимальная — 40,8 км/ч.
Дальность полета у различных особей пчелиной семьи (матки, пчел и трутней) различна. В открытой местности рабочие пчелы не улетают далее 4,8 км; в местности с неровным рельефом, пересеченной оврагами, покрытой деревьями дальность полета может достигать 14 км. Обычно радиус полета пчел с целью посещения источников пищи не превышает 3 км. Больше всего пчел семьи летает для сбора корма на расстояние 0,75 км от места размещения улья. За всю жизнь летная пчела налетывает, с целью сбора корма, до 800 км.
Матки с целью спаривания с трутнями улетают не далее 12 км от места размещения семьи. Нередко спаривание маток с трутнями происходит на расстоянии 5 км от улья. Считается, что оптимальная дальность полета маток на спаривание составляет около 2,5 км. Матки тратят на брачный полет в среднем около 20 мин.
Максимальное расстояние, на которое улетают трутни на спаривание, составляет около 7 км.
Высота полета пчел не превышает 10-12 м над зем-лей. Очень часто пчела при полете держится на высоте 1-1,5 м от земли.
Высота полета трутней и маток в зоне сбора в основном составляет 15-30 м, что облегчает их встречу. Максимальная высота нахождения трутней — 60 м. Во время полета к месту массового сбора трутни могут преодолевать горные преграды высотой до 1 км.
Брюшной отдел. У личинок всех трех особей пчелиной семьи брюшко состоит из 10 сегментов. В процессе превращения личинки во взрослую особь первое брюшное кольцо переходит в грудной отдел и образует проподеут. Последнее кольцо у всех трех особей образует анальное кольцо, которое внешне не обнаруживается.
Брюшко женских особей (пчелы и матки) состоит из шести хорошо видимых колец, 8 и 9 кольца пошли на образование жалоносного аппарата (рис. 12).
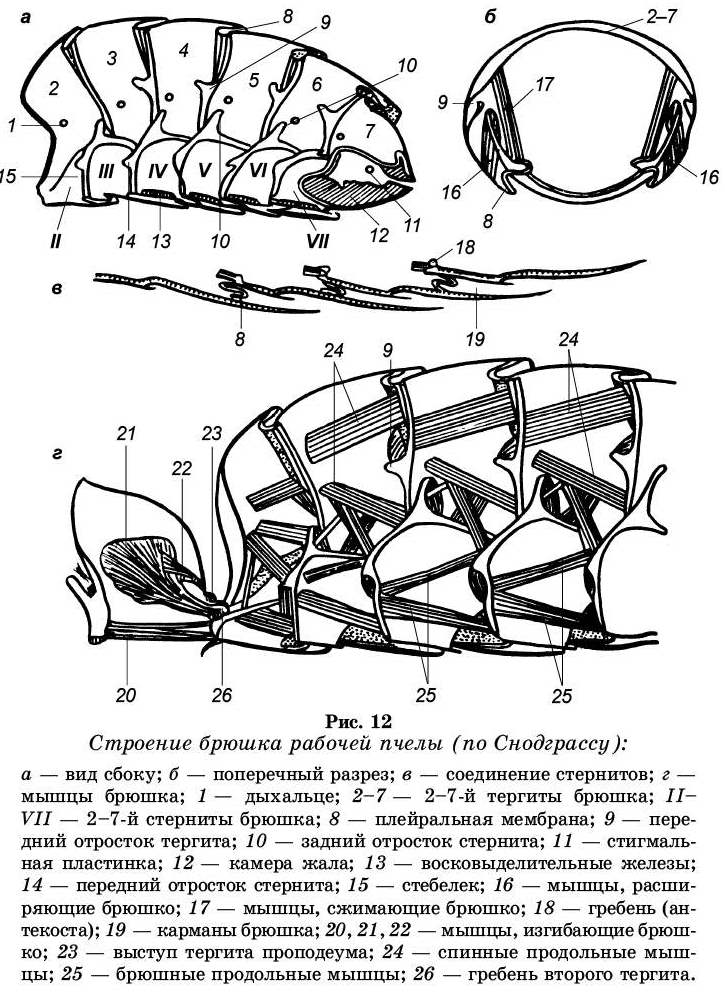
Брюшко трутня состоит из семи хорошо видимых колец. Девятое спинное полукольцо образует два выроста — молоточки, расположенные над половым отверстием, а брюшное его полукольцо срослось с полукольцом восьмого сегмента.
Каждое брюшное кольцо состоит из двух полуколец — спинного (тергита) и брюшного (стернита). Тергиты брюшка у пчелы сильно увеличены и прикрывают своими концами края стернитов. Концы тергитов и стернитов соединены между собой тонкой хитиновой перепонкой — плейральной мембраной.
Каждое брюшное кольцо соединяется с соседними также хитиновыми перепонками, причем предыдущее кольцо как бы прикрывает последующее кольцо. Такой способ соединения колец между собой и тергитов со стернитами обуславливает возможность расширения брюшка в продольном и поперечном направлениях. Способность брюшка к увеличению объема имеет огромное биологическое значение: в брюшке рабочей пчелы в основном сосредоточены органы пищеварения, а у матки и трутня значительное место занимают органы размножения. Постоянное изменение объема брюшка крайне необходимо для осуще-ствления процесса дыхания. Необходимость в значительном увеличении объема брюшка рабочей пчелы связана также со сбором корма, когда медовый зобик наполняется нектаром или при сборе воды, переносе меда. Увеличение объема брюшка крайне необходимо и во время зимовки, когда задняя кишка наполняется экскрементами. У маток после спаривания их с трутнями начинают интенсивно развиваться яйцевые трубочки яичников с развивающимися в них яйцеклетками, что также обеспечивается за счет уве-личения объема брюшка.
Тергит вдоль переднего края имеет утолщение — хитиновый валик с двумя бугорками. Каждый тергит имеет по бокам по паре дыхалец (стигм).
Стернит имеет хитиновый валик спереди и с боков. На валике переднего края имеется два бугорка, а на валике бокового края — отогнутые назад отростки.
Сегменты брюшка в значительной степени однотипны, отличаются они главным образом только величиной.
Первое брюшное кольцо спереди резко сужено в тонкий стебелек, соединяющий брюшко с грудью, обуславливающий его подвижность.
На четырех последних стернитах (брюшных полукольцах) рабочей пчелы имеется по два более тонких и светлоокрашенных участков хитина, называемые восковыми зеркальцами. На восковых зеркальцах в виде тонких пластинок образуется воск. С внутренней стороны восковые зеркальца выстланы железистыми клетками, которые выделяют воск.
Наружные края стернитов своими концами прикрывают начало стернитов последующих сегментов. В результате под каждым сегментом создаются как бы карманы, в которых помещаются выделившиеся восковые пластинки. В период интенсивного выделения воска у пчелы можно видеть края восьми восковых пластинок, выступающих из карманов.
У матки и трутня восковыделительные железы отсутствуют, и поэтому на стернитах нет восковых зеркалец. Стер-ниты матки более широкие. Стерниты трутня гораздо уже с глубокими перехватами по середине и с длинными игловидными боковыми отростками. Кроме того, девятый стернит недоразвит, но тергит этого сегмента сильно хитинизирован и имеет две пары пластинок, между которыми находится половое отверстие. При спаривании совокупительный орган трутня выворачивается наружу между этими пластинками.
Выросты тергитов и стернитов служат местом при-крепления мышц брюшка. Брюшные мышцы обеспечивают соединение тергитов со стернитами и соединяют отдельные членики между собой. Мускулатура брюшка представлена тремя группами:
■ первая — спинные продольные мышцы — проходят от
одной фрагмы к другой, соединяя членики;
■ вторая — брюшные продольные мышцы — соединяют
стерниты соседних сегментов;
■ третья дорсо-вентральные мышцы — соединяют тер-
гиты со стернитами.
Сокращение спинных и брюшных продольных мышц приводит к сближению сегментов и укорочению брюшка. За счет этих же групп мышц раздвигаются сегменты, что расширяет брюшко. Сокращение дорсо-вентральных мышц надвигает тергит на стернит, как бы сплющивает брюшко, что важно для дыхательных движений.
Жалоносный аппарат. Из восьмого и девятого брюшных сегментов у ряда насекомых образуется яйцеклад, а у жалоносных перепончатокрылых этот орган в процессе эволюционного развития видоизменился в жало.
Жало имеется у всех женских особей перепончатокрылых, однако его функции далеко не у всех одинаковы. У трутней жала нет.
У одних (пилильщики, наездники и др.) имеется яй-цеклад, функция которого заключается в том, что он прокалывает ткани растений или насекомого и откладывает туда яйца. У жалящих перепончатокрылых (пчелы, осы, муравьи) жало служит не только направляющим при откладывании яиц, но и органом защиты сообщества от нападения. При этом придаточные железы яйцекладущего аппарата превратились в производящие ядовитый секрет.
Брюшко пчелы состоит из шести сегментов. Последний сегмент брюшка конусообразный, внутри этого сегмента находится камера жала. Между тергитом и стерни-том этого сегмента сзади имеется щель, ведущая в камеру жала. Внутри камеры жала помещается жалоносный аппарат (рис. 13). Сверху жалоносный аппарат покрыт перепончатым тергитом восьмого брюшного сегмента. По бокам тергит склеротизирован и образует стигмальпые пла-
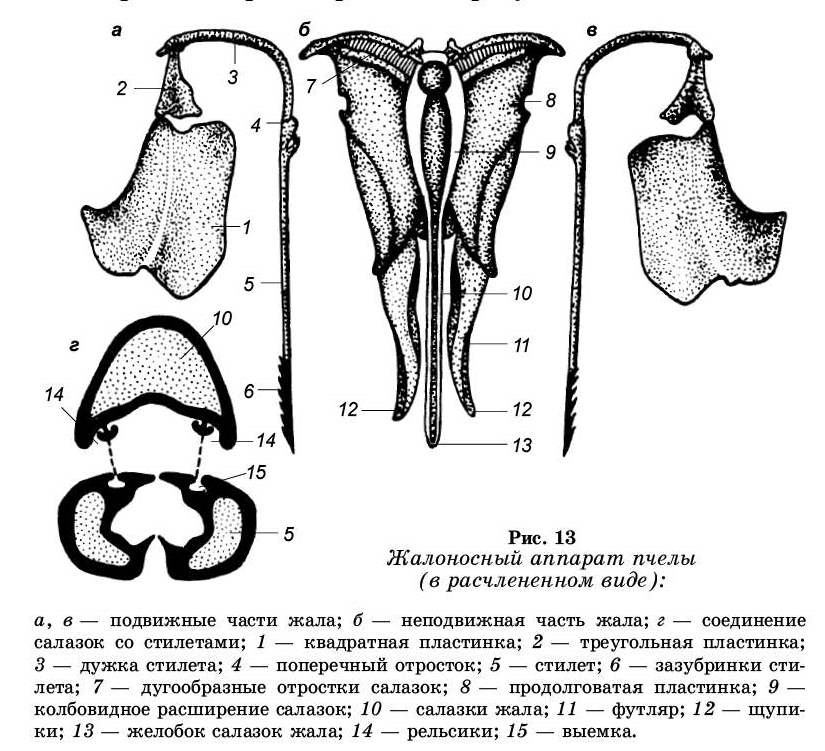
стинки. На каждой пластинке помещается дыхальце, или стигма. Все части жалоносного аппарата действуют как одно целое, несмотря на сложность его строения (в его состав входит более двадцати частей). Все составные части жала группируются вокруг двух отделов: подвижного и неподвижного.
Неподвижный отдел состоит из салазок. Салазки представляют собой длинный стержень, колбовидно расширенный в передней части и резко суженный в задней части. Суженная часть салазок имеет форму желобка, обращенного книзу. В колбовидном расширении салазок имеется большая полость, в которую впадают протоки двух ядовитых желез. Эта полость представляет собой вместилище для яда. От этой полости отходят узкий канал, по которому стекает яд. От колбовидного расширения салазок отходят в стороны и несколько вверх два плоских, дугообразных отростка — дуги салазок. Между ними находится вилочка, к которой прикрепляются мышцы.
К дугам салазок примыкают продолговатые пластинки. От заднего конца каждой продольной пластинки отходит назад мягкий футляр, переходящий на конце в щупик, усаженный чувствительными волосками.
Подвижный отдел — парное образование. Он состоит из стилетов — длинных тонких иголочек, имеющих полость внутри. На концах стилетов имеются 8-11 зазубри-нок, обращенных острием назад. У переднего конца стилеты загибаются в стороны, образуя дужки стилетов. Края дужек соединяются с углом треугольной пластинки, а треугольная пластинка соединяется с квадратной. У верхнего края квадратной пластинки находится смазочная железа, открывая профессором Г. А. Кожевниковым. Ее секрет служит для смазывания трущихся частей жалоносного аппа-рата. В месте перехода стилета в дужку находится хитиновый вырост — нагнетательный клапан.
Подвижная часть жала с неподвижной соединяются рельсообразно. Вдоль стилетов идут прорези в средней части, а вдоль салазок два выступа (валики салазок).
Жало матки по строению несколько отличается от жала рабочей пчелы: на стилетах всего 3-5 зазубринок, салазки жала имеют изогнутую форму.
С основанием салазок связаны ядовитые железы. Большая ядовитая железа рабочих пчел состоит из длинной тонкой трубочки с развилкой на конце и с резервуаром, в котором накапливается яд. Стенки железы выстланы слоем железистых клеток, выделяющих кислый секрет. Длина железы рабочих пчел из различных семей колеблется в широких пределах (от 9 до 20 мм). Установлено, что между длиной ядовитой железы и злобливостью пчел существует тесная прямая зависимость.
У матки большая ядовитая железа развита значительно сильнее, чем у рабочей пчелы. Длина ее составляет 40-50 мм. Раздваивается железа не на самом конце, а практически сразу же после отхода от резервуара. Сильное развитие большой ядовитой железы у матки, по сравнению с железой у рабочих пчел, объясняется тем, что она принимает участие в процессе кладки яиц. На поверхности откладываемых маткой яиц всегда содержится тонкий слой секрета этой железы. Известно, что свежевыде-ленный яд стерилен и обладает сильнейшими бактерицидными свойствами.
Малая ядовитая (дюффуровская) железа представлена в виде короткой извилистой трубочки, не имеющей резервуара. Выводной проток ее расположен рядом с выводным протоком большой ядовитой железы. Секрет железы имеет щелочную реакцию, по этой причине ее еще называют щелочной железой. Отдельно секреты этих желез ядовитых свойств не имеют. Ядовитые свойства приобретаются при смешивании секретов, которые происходят в полости колбовидного расширения салазок, куда впадают протоки желез.
Молодые пчелы, только что вышедшие из ячеек, запаса яда в резервуаре вовсе не имеют. В отличие от рабочих пчел, матки выходят из ячейки, уже имея в резервуаре ядовитый секрет, который используется для борьбы с другими активными матками. Со 2-3 дня жизни, когда пчелы начинают потреблять большое количество белкового корма, секреторные клетки желез начинают развиваться и продуцировать яд. К 11-16-суточному возрасту (в среднем к 12-му дню их жизни) в резервуаре железы накапливается максимальное количество яда, после чего секреторные клетки дегенерируют. У 12-дневных пчел сухая масса ядовитой железы и ядосборника резко повышается и достигает максимальной величины за весь период жизни пчел. У пчел старше 14-20 дней способность образования яда прекращается. Ко времени перехода пчел к полевым работам и выполнению функций по охране гнезда деятельность желез полностью прекращается. Готовый яд все время хранится в резервуаре железы. В большинстве случаев сторожевая пчела погибает после ужаления, израсходовав сразу весь запас яда. В этом и состоит целесообразность атрофии железистых клеток после того, как резервуар ядовитой железы полностью заполнится. Израсходованный запас яда у пчел не восстанавливается.
У летних пчел (июльских) ядовитая железа и резервуар развиты лучше, чем у весенних (майских) и осенних (сентябрьских) пчел, и соответственно этому изменяется и количество яда в резервуаре. У июньских и июльских пчел наивысшая активность яда.
У пчел осенней генерации секреторная деятельность клеток ядовитых желез начинается впервые лишь с 14-15 дня жизни, а к 20-му дню их деятельность прекращается. Осеннее поколение пчел имеет достоверно меньше яда в резервуаре желез, чем пчелы весенне-летних генераций.
Качественный состав яда также зависит от возраста пчел. Так, главный компонент яда мелиттин в резервуаре рабочих пчел 2-3-дневного возраста представлен неактивной формой — промелиттином, и активизируется максимально к 20-му дню их жизни. У матки превращение промелиттина в мелиттин происходит уже на 1-й день жизни. Гистамин и гистидин имеют максимальное содержание также к 20-му дню жизни рабочей пчелы.
Кроме возраста пчел, времени года на количество яда также влияет порода пчел. Пчелы среднерусской породы в резервуаре ядовитой железы имеют в среднем 0,43 + + 0,040 мг пчелиного яда (с разбросом от 0,180 до 0,760 мг), а у пчел серой горной кавказской породы — 0,36 + 0,023 мг (с разбросом от 0,190 до 0,580 мг). В резервуаре железы одной пчелы краинской породы находится количество яда, среднее между пчелами среднерусской и серой горной кавказской пород (в среднем 0,30 ± 0,017 мг).
В расчете на сухой яд от одной пчелы среднерусской породы можно получить в среднем около 0,146 ± 0,030 мг, а от пчелы серой горной кавказской породы 0,124 ± 0,018 мг яда-сырца. Таким образом, от одного килограмма рабочих пчел (в среднем 10 000 особей) можно практически получить в среднем 1,0 г яда-сырца.
Наконец, на количество яда в резервуаре железы пчелы значительное влияние оказывает сила семей и обеспеченность их полноценными обильными кормами и особенно белковыми. Полноценное белковое питание молодых пчел во многом определяет уровень развития секреторных клеток ядовитых желез, количество яда и его биохимический состав (качество). Установлено, что пчелы, выкормленные без пыльцы, вовсе не вырабатывают яда.
Жаление. Жалоносный аппарат у спокойной пчелы полностью спрятан внутри последнего сегмента брюшка, в камере жала. Перед ужалением аппарат выводится наружу (стилетами вперед) за счет сокращения мышц передней части брюшка. Жало обнажается, и концы его стилетов выдвигаются далеко наружу, на которых выступает первая капелька яда.
Жалоносный аппарат соединен с последним сегментом брюшка таким образом, что после ужаления он легко отрывается. Жалоносный аппарат, будучи оторванным от тела пчелы, и, находясь в ранке животного, функционирует самостоятельно, вследствие чего происходит дальнейшее внедрение в ранку все глубже и глубже. При этом поперечные отростки стилетов при каждом движении нагнетают в ранку все новые порции яда. На изливание всего яда наружу требуется несколько минут. Отсюда следует, что для полного извлечения яда необходимо, чтобы жалоносный аппарат был оторван от тела пчелы. Автоматизм действия жалоносного аппарата объясняется тем, что одновременно с ним отрывается последний нервный ганглий, который вместе с мускулатурой обеспечивает работу жала, оказавшегося вне тела пчелы. Отрыв жала приводит к гибели пчелы.
Легкий отрыв жала является важным приспособлением, повышающим эффективность жаления в борьбе с врагами, разоряющими гнезда пчел.
Пчела может защищаться самостоятельно, а также мобилизовать на помощь других членов семьи — своих сестер. В мобилизации пчел на защиту своего гнезда важная роль принадлежит пахучим веществам, выделяемым возбужденными пчелами, которые выполняют роль сигналов тревоги. Хорошо известно, что яд, выделяемый возбужденными пчелами в момент ужаления, обладает сильным запахом. Считалось, что он приводит в возбуждение пчел и служит сигналом для защиты гнезда от врагов. В настоящее время установлено, что в самом яде нет веществ, возбуждающих пчел и служащих для них сигналом тревоги. Феромон тревоги выделяется железами, расположенными у основания жала. Из жала рабочих пчел выделен изоамилацетат. Он вызывает агрессивные реакции особей, подвергаемых раздражению этим веществом. Приводит в возбуждение с характерной агрессивной реакцией (пчелы принимают специфическую позу: поднимают вверх брюшко, выпячивают жало, на кончике которого появляется капелька яда, и машут крыльями) также гектан-2-он, выделяемый мандибулярными железами.
В том случае, когда пчеле удается после ужаления вытянуть жало обратно, она остается жить. Хотя известно, что после максимальной отдачи яда пчела становится менее активной, с пониженной способностью к защите своего гнезда. Как правило, пчела, которая не потеряла жала, не отдает полный запас своего яда.
Химический состав, биохимические и физические свой-ства пчелиного яда. Свежесекретированный яд представляет собой прозрачную бесцветную жидкость со специфическим запахом и горько-жгучим вкусом, с кислой реакцией (рН 4,5-5,5), плотностью 1,08-1,13, быстровысыхающую на воздухе. Летучие вещества пчелиного яда составляют вода и сложные эфиры: изоамилацетат, изоамилпропинат, изоамил-бутират и др. Сухой пчелиный яд представляет собой порошок в виде чешуек и крупинок серовато-желтого (до бурого) цвета, масса которого составляет 30-40% от нативного секрета. Коричневатый цвет свидетельствует о фотоокислении на солнце триптофана боковых цепочек в белках яда.
Яд растворяется в воде, кислотах и не растворяется в спирте, разрушается в концентрированных кислотах и щелочах, этиловом спирте, а также под действием солнечного света и высоких температур.
В пчелином яде содержатся ферменты, пептиды, аминокислоты, жиры и стерины, глюкоза, фруктоза, жирные кислоты, зольные элементы — всего 50 веществ. Основная часть сухого вещества яда представлена белками и пептидами — около 80°/). Все они являются активными веществами. Основным компонентом яда (около 50°/) сухого вещества) является белок мелиттин (см. табл. 1).
Гиалуронидазе принадлежит функция выработки иммунитета организма. Этот фермент гидролизует вязкую гиалуроновую кислоту на невязкие компоненты. Гиалуро-новая кислота способствует удержанию клеток тканей организма вместе, а при ее разрушении гиалуронидазой промежутки между клетками становятся невязкими, что облегчает проникновение других составляющих яда. Ее роль, таким образом, состоит в содействии проникновения пчелиного яда в организм, она способствует также рассасыванию гема-том, шрамов и прочих затвердений соединительной ткани. Фосфолипаза А2, проникая в организм, усиливает воспалительный процесс, вызываемый действием яда, снижает активность антигенов. Она разрушает главный строительный блок всех биохимических мембран — фосфолипиды. Фосфолипаза пчелиного яда — самая активная из всех известных фосфолипаз, она активнее фосфолипаз змеиного яда, а также панкреатической фосфолипазы млекопитающих.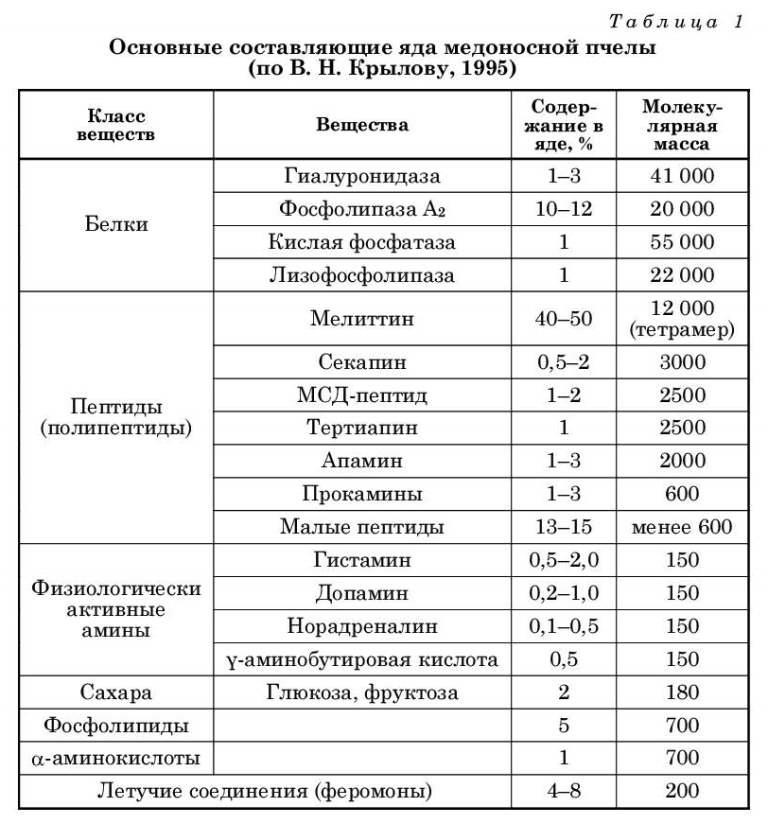
Токсичность и терапевтическое действие яда определяются, в основном, мелиттином и, отчасти, апамином.
Мелиттин, проникая в организм, вызывает гемолиз эритроцитов и спазмы мышц, уменьшает свертываемость крови, обладает противомикробным действием. Действие мелиттина зависит от дозировки. Мелиттин проявляет также лучезащитное действие. В больших дозах этот пептид токсичен. Мелиттин является слабым аллергеном.
Мелиттин и фосфолипаза токсичны сами по себе, но при совместном действии их токсичность сильно возрастает.
Апамин возбуждающе действует на нервную систему, он токсичен, активизирует функцию желез внутренней секреции, повышает кровяное давление. Он блокирует каналы мембран и ингибирует действие адреналина, который должен открывать эти каналы.
Яд пчел резко отличается от всех остальных ядов: от яда гадюки отличается тем, что является антикоагулянтом, в то время как пчелиный — коагулянт; от яда кобры он отличается содержанием в нем факторов диффузии.
Энзимы пчелиного яда в 30 раз активнее, чем змеиного.
Действие пчелиного яда на организм. Общее действие яда сводится к нейротоксическому, геморрагическому и гемолитическому. Под влиянием различных доз яда отмечается падение кровяного давления, изменение ритма сердцебиений с возникновением аритмий, затруднение дыхания из-за спазмов гладкой мускулатуры воздухопроводя-щих путей и угнетения дыхательного центра.
При ужалении человек ощущает довольно сильную, но непродолжительную боль. Через несколько часов вокруг ужаленного места возникает краснота и припухлость. Обычно местная реакция держится несколько часов или суток, после чего проходит бесследно. Прекращается болевое ощущение, затем проходит краснота и спадает отек.
Чувствительность разных животных к пчелиному яду различна. Для всех насекомых одно ужаление пчелы смертельно. Мыши погибают от 2-3 ужалении, цыплята — от 3-5, куры — от 8-10, для лошади смертельна доза свыше 1000 ужалении. Чувствительность организма человека к пчелиному яду сильно различается. Встречаются люди, у которых повышенная чувствительность к пчелиному яду, и уже одно ужаление вызывает у них признаки, напомина-ющие отравление. Напротив, есть люди, организм которых
практически не чувствителен к пчелиному яду. Обычно 1-10 ужалений одновременно человек переносит без проявлений общих симптомов, отвечая лишь местной реакцией. При 200-400 ужалениях люди уже серьезно поражаются ядом с развитием симптомов общей интоксикации. Считается, что 500 ужалений одновременно приводит к смертельному исходу. Пчеловоды, в организм которых длительное время в небольших дозах постоянно поступает пчелиный яд, становятся к нему невосприимчивы, наступает так называемый митридатизм.
Малые дозы пчелиного яда с давних пор используют при лечении различного рода сомато-вегетативных нарушений. В Китае в настоящее время лечат пчелиным ядом более 400 заболеваний. Пчелиным ядом лечат остеохондрозы и невриты, невралгии и полиневриты, импотенцию и фригидность, полиартриты и почечную недостаточность, гинекологические и хирургические заболевания. В лечении основное значение имеет повышение защитных сил организма, получающего яд. Перспективным направлением применения пчелиного яда является его радиопротекторное и иммуномобилизирующее действие.